




Articles
The Protein Debate: Dr. Loren Cordain & T. Colin Campbell
March 19, 2008

March 19, 2008
Introduction
Protein plays a litany of roles in living systems: structural elements, peptide hormones, cell recognition, antibodies… the list is staggering and continues to grow as our understanding of biology expands. What, however, is the role of dietary protein in health and disease in humans? Is the source, type and quantity intimately and directly tied to optimal physical development and continued wellbeing? Is it causative or preventative of disease? How do we know, and how can we know?
One would think this question should be straightforward and easily answered; as you will soon see the question is anything but simple! In the pages that follow, two scientists at the top of their respective fields--Dr. T. Colin Campbell, Professor of Nutritional Biochemistry at Cornell University, author of The China Study and Dr. Loren Cordain Professor, Department of Health & Exercise Science, Colorado State University, author of The Paleo Diet—make their competing cases for the role of dietary protein in health and disease.
The Evolutionary Basis for the Therapeutic Effects of High Protein Diets
Loren Cordain, Ph.D, Professor
Department of Health and Exercise Science
Colorado State University
Introduction
Although humanity has been interested in diet and health for thousands of years, the organized, scientific study of nutrition has a relatively recent past. For instance, the world’s first scientific journal devoted entirely to diet and nutrition, The Journal of Nutrition only began publication in 1928. Other well known nutrition journals have a more recent history still: The British Journal of Nutrition (1947), The American Journal of Clinical Nutrition (1954), and The European Journal of Clinical Nutrition (1988). The first vitamin was “discovered” in 1912 and the last vitamin (B12) was identified in 1948 (1). The scientific notion that omega 3 fatty acids have beneficial health effects dates back only to the late 1970’s (2), and the characterization of the glycemic index of foods only began in 1981 (3).
Nutritional science is not only a newly established discipline, but it is also a highly fractionated, contentious field with constantly changing viewpoints on both major and minor issues that impact public health. For example, in 1996 a task force of experts from the American Society for Clinical Nutrition (ASCN) and the American Institute of Nutrition (AIN) came out with an official position paper on trans fatty acids stating, “We cannot conclude that the intake of trans fatty acids is a risk factor for coronary heart disease” (4).
Fast forward 6 short years to 2002 and the National Academy of Sciences, Institute of Medicine’s report on trans fatty acids (5) stating, “Because there is a positive linear trend between trans fatty acid intake and total and LDL (“bad”) cholesterol concentration, and therefore increased risk of cardiovascular heart disease, the Food and Nutrition Board recommends that trans fatty acid consumption be as low as possible while consuming a nutritionally adequate diet”.
These kinds of complete turnabouts and divergence of opinion regarding diet and health are commonplace in the scientific, governmental and medical communities. The official U.S. governmental recommendations for healthy eating are outlined in the “My Pyramid” program (6) which recently replaced the “Food Pyramid”, both of which have been loudly condemned for nutritional shortcomings by scientists from the Harvard School of Public Health (7). Dietary advice by the American Heart Association (AHA) to reduce the risk of coronary heart disease (CHD) is to limit total fat intake to 30% of total energy, to limit saturated fat to <10% of energy and cholesterol to <300 mg/day while eating at least 2 servings of fish per week (8). Although similar recommendations are proffered in the USDA “My Pyramid”, weekly fish consumption is not recommended because the authors of these guidelines feel there is only “limited” information regarding the role of omega 3 fatty acids in preventing cardiovascular disease (6). Surprisingly, the personnel makeup of both scientific advisory boards is almost identical. At least 30 million Americans have followed Dr. Atkins advice to eat more fat and meat to lose weight (9). In utter contrast, Dean Ornish tells us fat and meat cause cancer, heart disease and obesity, and that we would all would be a lot healthier if we were strict vegetarians (10). Who’s right and who’s wrong? How in the world can anyone make any sense out of this apparent disarray of conflicting facts, opinions and ideas?
In mature and well-developed scientific disciplines there are universal paradigms that guide scientists to fruitful end points as they design their experiments and hypotheses. For instance, in cosmology (the study of the universe) the guiding paradigm is the “Big Bang” concept showing that the universe began with an enormous explosion and has been expanding ever since. In geology, the “Continental Drift” model established that all of the current continents at one time formed a continuous landmass that eventually drifted apart to form the present-day continents. These central concepts are not theories for each discipline, but rather are indisputable facts that serve as orientation points for all other inquiry within each discipline. Scientists do not know everything about the nature of the universe, but it is absolutely unquestionable that it has been and is expanding. This central knowledge then serves as a guiding template that allows scientists to make much more accurate and informed hypotheses about factors yet to be discovered.
The study of human nutrition remains an immature science because it lacks a universally acknowledged unifying paradigm (11). Without an overarching and guiding template, it is not surprising that there is such seeming chaos, disagreement and confusion in the discipline. The renowned Russian geneticist Theodosius Dobzhansky (1900-1975) said, “Nothing in biology makes sense except in the light of evolution” (12). Indeed, nothing in nutrition seems to make sense because most nutritionists have little or no formal training in evolutionary theory, much less human evolution. Nutritionists face the same problem as anyone who is not using an evolutionary model to evaluate biology: fragmented information and no coherent way to interpret the data.
All human nutritional requirements like those of all living organisms are ultimately genetically determined. Most nutritionists are aware of this basic concept; what they have little appreciation for is the process (natural selection) which uniquely shaped our species’ nutritional requirements. By carefully examining the ancient environment under which our genome arose, it is possible to gain insight into our present day nutritional requirements and the range of foods and diets to which we are genetically adapted via natural selection (13-16). This insight can then be employed as a template to organize and make sense out of experimental and epidemiological studies of human biology and nutrition (11).
The Dietary Protein Conundrum: How Much is Enough?
An important dietary issue that has come under debate in recent years is the safety of high protein diets and their long term influence upon health and well being (17, 18). In the current U.S. diet the average protein intake is 98.6 g/day (15.5 % of total energy) for men and 67.5 g/day (15.1 % of total energy) for women (19). Animal products provide approximately 75 % of the protein in the U.S. food supply followed by dairy, cereals, eggs, legumes, fruits and vegetables (20). Diets containing 20 % or more of their total energy as protein have been labeled “high protein diets” and those containing 30% or more energy as protein have been dubbed “very high protein diets” (18). Accordingly, a “high protein diet” for the average U.S. male daily energy intake (2,618 kcal (19)) would contain between 125 to 186 grams of protein per day and for the average female (1,877 kcal (19)) between 89 to 133 grams of protein per day.
At this point, it should be noted that there is a physiological limit to the amount of protein that can be ingested before it becomes toxic (14, 21). A byproduct of dietary protein metabolism is nitrogen, which in turn is converted into urea by the liver and then excreted by the kidneys into the urine. The upper limit of protein ingestion is determined by the liver’s ability to synthesize urea. When nitrogen intake from dietary protein exceeds the ability of the liver to synthesize urea, excessive nitrogen (as ammonia) spills into the bloodstream causing hyperammonemia and toxicity (14, 21). Additionally excess amino acids from the metabolism of high amounts of dietary protein may become toxic by entering the circulation causing hyperaminoacidemia (14, 21).
The avoidance of the physiological effects of protein excess has been an important factor in shaping the subsistence strategies of hunter-gatherers (22- 24). Multiple historical and ethnographic accounts have documented the deleterious health effects that have occurred when humans were forced to rely solely upon the fat depleted, lean meat of wild animals (22). Excess consumption of dietary protein from the lean meats of wild animals leads to a condition referred to by early American explorers as “rabbit starvation” which initially results in nausea, then diarrhea and eventual death (22). Clinical documentation of this syndrome is virtually non-existent, except for a single case study (25).
Using known maximal rates of urea synthesis (MRUS) in normal subjects [65 mg N/h - kg (body weight )0.75 ] (range 55-76), it is possible to calculate the maximal protein intake, beyond which will exceed MRUS and result in hyperammonemia and hyperaminoacidemia (21). The mean maximal protein intake for the average weight U.S. male (189.4 lbs (26)) is then 270 g/day (range 233-322 g/day), and for an average weight female (162.8 lbs (26)), 246 g/day (range 208-288 g/day). Consequently, “very high protein diets” for the average U.S. male could range from 187 to 270 g/day and for females, 134 to 246 g/day.
So let’s summarize a few key points. The average protein intake in the U.S. is about 15 % of the normal daily caloric intake. Diets labeled as “high protein” contain 20-29 % protein of the normal daily caloric intake, and diets with 30-40 % protein are branded “very high protein”. It should be pointed out that this categorization is completely arbitrary and based almost entirely upon comparisons to the U.S. norm. A salient question from an evolutionary perspective would be, “Is the average U.S. protein intake necessarily average or normal for our species?” For example, blood pressure in the U.S. and most other westernized countries is considered “normal” when systolic pressure is 120 mm Hg and diastolic pressure is 80 mm Hg. However, in many non-westernized people these values would be higher than normal. Consider the data in Figure 1 below showing blood pressure in the Yanomamo Indians of Brazil, a non-salt consuming society. Not only is blood pressure lower than normal western values, but it stays uniform throughout life and does not rise with age (27).
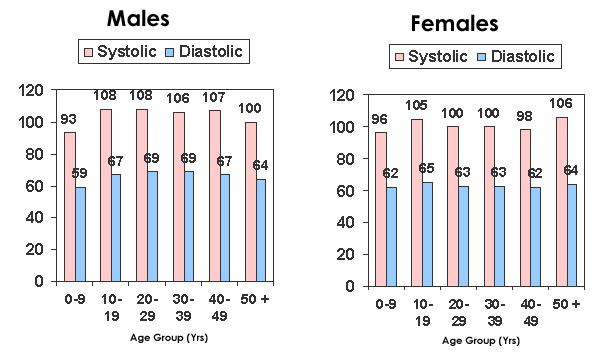
Figure 1. Blood pressure in a group of 506 Brazilian Indians (26).
In order to objectively answer the question whether or not high protein diets have detrimental or therapeutic health effects compared to the U.S. norm (15 % total energy), it may be useful to frame this question in an evolutionary perspective before examining the experimental and epidemiological evidence.
High Protein Diets: The Evolutionary Evidence
The Fossil Evidence
A number of lines of evidence suggest that meat eating and high protein diets have been a component of human nutrition since the very origins of our genus Homo. Beginning approximately 2.6 million years ago (MYA), the hominin species that eventually led to Homo began to include more animal food in their diet. A number of lines of evidence support this viewpoint. First, the very first stone tools (Oldowan lithic technology) appear in the fossil record 2.6 MYA (28), and there is clear cut evidence to show that these tools were used to butcher and disarticulate animal carcasses (29, 30). Stone tool cut marks on the bones of prey animals and evidence for marrow extraction appear concurrently in the fossil record with the development of Oldowan lithic technology by at least 2.5 MYA (Figure 2) (30). It is not entirely clear which specific early hominin species or group of species manufactured and used these earliest of stone tools; however, Australopithecus garhi is a likely candidate (30, 31).
The development of stone tools and the increased dietary reliance on animal foods allowed early African hominins to colonize northern latitudes outside of Africa where plant foods would have been seasonally restricted. Early Homo skeletal remains and Oldowan lithic technology appear at the Dmanisi site in the Republic of Georgia (40° N) by 1.75 MYA (32), and more recently Oldowan tools dating to 1.66 MYA have been discovered at the Majuangou site in North China (40° N) (33). Both of these tool-producing hominins would likely have consumed considerably more animal food than pre-lithic hominins living in more temperate African climates, and it is likely the majority of their daily energy was obtained from animal foods during winter and early spring when plant food sources would have been scarce or unavailable.
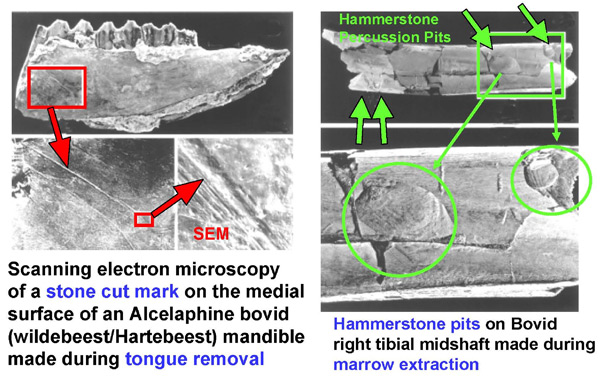
Figure 2: The earliest evidence for meat and marrow extraction dating to 2.5 million years ago (30).
The Genetic Evidence
In addition to the fossil evidence suggesting a trend for increased animal food consumption, hominins may have experienced a number of genetic adaptations to animal-based diets early in our genus’s evolution analogous to those of obligate carnivores such as felines. Carnivorous diets reduce evolutionary selective pressures that act to maintain certain anatomical and physiological characteristics needed to process and metabolize high amounts of plant foods. In this regard, hominins, like felines, have experienced a reduction in gut size and metabolic activity along with a concurrent expansion of brain size and metabolic activity as they included more energetically dense animal food into their diets (16, 34, 35). Further, similar to obligate carnivores (36), humans maintain an inefficient ability to chain elongate and desaturate 18 carbon fatty acids to their product 20 and 22 carbon fatty acids (37). Since 20 and 22 carbon fatty acids are essential cellular lipids, then evolutionary reductions in desaturase and elongase activity in hominins indicate that preformed dietary 20 and 22 carbon fatty acids (found only in animal foods) were increasingly incorporated in lieu of their endogenously synthesized counterparts derived from 18 carbon plant fatty acids. Finally, our species has a limited ability to synthesize the biologically important amino acid, taurine, from precursor amino acids (38, 39), and vegetarian diets in humans result in lowered plasma and urinary concentrations of taurine (40). Like felines (41, 42) the need to endogenously synthesize taurine may have been evolutionarily reduced in humans because exogenous dietary sources of preformed taurine (found only in animal food) had relaxed the selective pressure formerly requiring the need to synthesize this conditionally essential amino acid.
Another genetic adaptation to a high meat diet involves the metabolism of purines. Purines are the nitrogenous base pairs which form the structural cross rung molecules of both DNA and RNA. As DNA and RNA are broken down within cells, the purines then can be metabolized into uric acid by the liver and a few other tissues within the body. The liver receives purines from two sources: 1) the diet, and 2) the daily breakdown of the body’s own tissues. About 2/3 of the daily purine load comes from the body’s turnover of cells, while 1/3 comes from the diet (43). When the combined purine load (from both diet and turnover of the body’s own cells) exceeds the kidney’s ability to excrete it, blood concentrations of uric acid rise, thereby increasing the risk for gout, a painful disease caused by formation of uric acid crystals in the joints. Although high protein, meat based diets contain high amounts of purines and would be expected to promote gout symptoms, protein ingestion actually decreases blood uric acid levels by increasing uric acid excretion (44). This seemingly paradoxical effect occurs because the kidney increases its excretion of uric acid when faced with elevated dietary purines (45). But more importantly, over the course of evolution, humans have evolved a genetic mutation which tends to prevent uric acid synthesis in the liver. Humans avoid the overproduction of uric acid in the face of increasing dietary purine intake from meats by decreasing the activity of an enzyme called xanthine oxidoreductase (46), a key catalyst in the final synthesis of uric acid. Compared to other animals, xanthine oxidase activity is almost 100 times lower in humans (47). This evolutionary adaptation has occurred because the gene coding for xanthine oxidoreductase has been repressed (48). The final proof of the pudding has been borne out by dietary interventions showing that high protein, low glycemic load diets actually normalized serum uric acid concentrations in 7 of 12 gout patients and significantly decreased gout attacks (49).
The Isotopic Fossil Evidence
Since the evolutionary split between hominins and pongids (apes) approximately 7 million years ago, the available evidence shows that all species of hominins ate an omnivorous diet composed of minimally processed, wild-plant, and animal foods. In support of this view is the omnivorous nature of chimpanzees, the closest living pongid link to hominins. Although chimpanzees (Pan paniscus and Pan troglodytes), our genetically closest nonhuman relatives, primarily consume a frugivorous diet, they still eat a substantial amount of meat obtained throughout the year from hunting and scavenging (50-52). Observational studies of wild chimpanzees demonstrate that during the dry season meat intake is about 65 g per day for adults (51). Accordingly, it is likely that the very earliest hominins would have been capable of obtaining animal food through hunting and scavenging in a manner similar to chimpanzees.
Carbon isotope data also support the notion that early hominins were omnivorous. By about 3 million years ago MYA Australopithecus africanus obtained a significant portion of food from C4 sources (grasses, particularly seeds and rhizomes; sedges; invertebrates, including locusts and termites; grazing mammals; and perhaps even insectivores and carnivores)(53). Other fossils of early African hominins, including Australopithecus robustus and Homo ergaster, maintain carbon isotope signatures characteristic of omnivores (54, 55). The finding of C4 in Australopithecus robustus fossils refutes the earlier view that this hominin was vegetarian (54).
There is little evidence to the contrary that animal foods have always played a significant role in the diets of all hominin species. Increased reliance on animal foods not only allowed for enhanced encephalization (brain expansion relative to body weight) and its concomitant behavioral sophistication (16, 34, 35), but this dietary practice also permitted colonization of the world outside of Africa. An unresolved issue surrounding hominin diets is the relative amounts of plant and animal foods that were typically consumed.
Before the advent of Oldowan lithic technology about 2.6 MYA quantitative estimates of hominin energy intake from animal food sources are unclear, other than they were likely similar to, or greater than, estimated values (4%–8.5% total energy) for chimpanzees (51, 56)). Although all available data point to increasing animal food consumption following the arrival of stone tool technology, the precise contribution of either animal or plant food to energy is unclear. Obviously, then as now, no single (animal/plant) subsistence ratio would have been necessarily representative of all populations or species of hominins. However, there are a number of lines of evidence which suggest more than half (>50%) of the average daily energy intake for most Paleolithic hominin species and populations of species was obtained from animal foods.
Richards, Pettitt, and colleagues (57) have examined stable isotopes (δ13C and δ15N) in two Neanderthal specimens (~28,000—29,000 years ago) from Vindija Cave in northern Croatia and contrasted these isotopic signatures to those in fossils of herbivorous and carnivorous mammals from the same ecosystem. The analysis demonstrated that Neanderthals, similar to wolves and arctic foxes, behaved as top-level carnivores, obtaining all of their protein from animal sources (57). More recent studies corroborate this earlier work and points to Neanderthals “as top predators in an open environment, with little variation through time and space” (58), and “the percentage of plants in the Neanderthal diet must have been close to zero (59).
Because Neanderthals were not direct predecessors of modern humans (60), it may be more relevant to examine the isotopic data from fully modern humans living during the Pleistocene. An analysis was made of five Upper Paleolithic Homo sapiens specimens dated to ~11,700–12,380 years ago from Gough’s and Sun Hole Caves in Britain (61). The data indicated these hunter-gatherers were consuming animal protein year-round at a higher trophic level than the artic fox.
All of these studies (57-62) could be criticized as not being representative of typical hominin diets, as these two species lived in climates and ecosystems that fostered an abundance of large, huntable mammals, which were preyed upon preferentially. Additional clues to the typical plant-to-animal subsistence ratio in Paleolithic hominin diets can be found in the foraging practices of historically studied hunter-gatherers.
The Ethnographic Evidence
Our analysis (Figure 3) of the Ethnographic Atlas data (62) showed that the dominant foods in the majority of historically studied hunter-gatherer diets were derived from animal food sources (14). Most (73%) of the world’s hunters-gatherers obtained >50 percent of their subsistence from hunted and fished animal foods, whereas only 14 percent of worldwide hunter gatherers obtained >50 percent of their subsistence from gathered plant foods. For all 229 hunter-gatherer societies, the median subsistence dependence on animal foods was 56 percent to 65 percent. In contrast, the median subsistence dependence on gathered plant foods was 26 percent to 35 percent (14).
The major limitation of ethnographic data is that the preponderance of it is subjective in nature, and the assigned scores for the five basic subsistence economies in the Ethnographic Atlas are not precise, but rather are approximations (63). Fortunately, more exact, quantitative dietary studies were carried out on a small percentage of the world’s hunter gatherer societies (15, 64). Table 1 lists these studies and shows the plant-to-animal subsistence ratios by energy. The average score for animal food subsistence is 65 percent, while that for plant-food subsistence is 35 percent. These values are similar to our analysis of the entire (n = 229) sample of hunter-gatherer societies listed in the Ethnographic Atlas in which the mean score for animal food subsistence was 68 percent and that for plant food was 32 percent (14). When the two polar hunter-gatherer populations, who have no choice but to eat animal food because of the inaccessibility of plant foods, are excluded from Table 1 the mean score for animal subsistence is 59 percent and that for plant-food subsistence is 41 percent. These animal-to-plant subsistence values fall within the same respective class intervals (56%–65% for animal food; 26%–35% for plant food) as those we estimated from the ethnographic data when the confounding influence of latitude was eliminated (14). Consequently, there is remarkably close agreement between the quantitative data in Table 1 and the ethnographic data (14) that animal food comprised more than half of the energy in historically studied hunter-gatherer diets.
Based upon hunter gatherer plant to animal subsistence ratios and the known macronutrient contents of wild plant and animal foods, it is possible to estimate the macronutrient content of these diets (14). The typical hunter-gatherer protein intake would have fallen between 19 and 35 % of total energy (14), values which would be labeled either “high” or “very high” protein diets when compared to current U.S. values (15 %). Consequently, when framed in an evolutionary context, current western dietary protein intakes fall outside the range of diets that would have conditioned the human genome for nearly 2 million years. The evolutionary template would then suggest that when dietary protein intakes are restored to levels that our species is genetically accustomed, good health will prevail. Conversely, lower or higher values likely result in ill health. Let’s see what the experimental evidence shows.
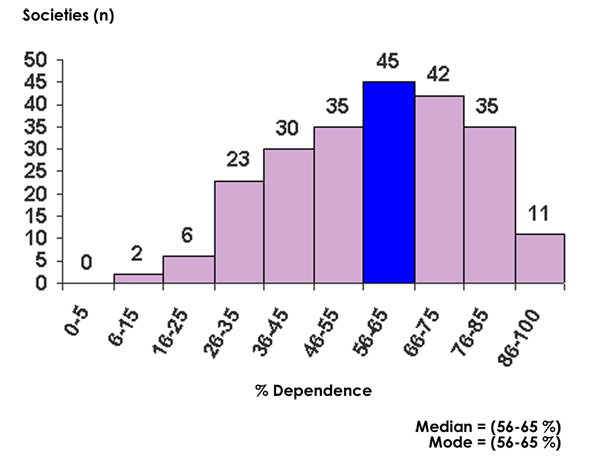
Figure 3. Frequency distribution of subsistence dependence upon animal foods in world wide hunter gatherer societies (n = 229).
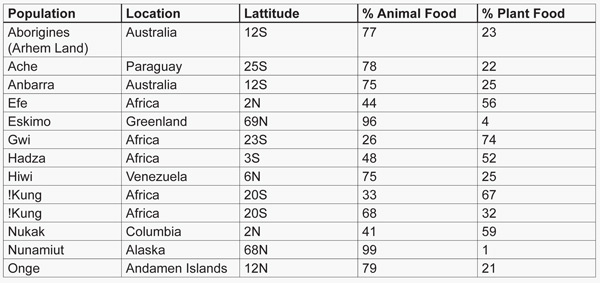
Table 1. Quantitatively determined proportions of plant and animal food in hunter-gatherer diets (15, 64).
Dietary Protein: Health and Well Being
Establishing Cause and Effect between Diet and Disease
One of the challenges faced by nutritional scientists when they ultimately make recommendations regarding what we should and should not eat is to establish cause and effect between a dietary element and the subsequent development or prevention of disease. Some foods and some dietary habits promote good health whereas others promote disease. Figure 4 demonstrates the four primary procedures by which causality is established between diet and disease (65, 66).
No single procedure alone can establish cause and effect (65, 66), nor can any single study prove causality (67). Observational epidemiological studies can only show relationships among variables and are notorious for showing conflicting results (68 ) and cannot provide decisive evidence by themselves either for or against specific hypotheses (69). For example increased animal protein has been associated with a decreased risk for coronary heart disease (CHD) in a large group of nurses (The Nurses Health Study) (70), whereas exactly the opposite association was found for markers of CHD and meat consumption in people from rural China (71, 72). An analogy here may be appropriate to show you why observational epidemiological studies can only show relationships and not establish causality. In New York City, there is a strong association between the size of a structure fire and the number of fire trucks at the fire, but can we conclude that more fire trucks cause bigger fires?
In order to establish cause and effect between diet and disease, it takes more than just observational epidemiological evidence (69). There must also be what is referred to as “biological plausibility” in which evidence gathered from tissue, animal and short term human metabolic studies support causality (66). When observational epidemiological evidence is augmented by biological plausibility studies and confirmed by randomized controlled trials, the case for causality becomes ever more convincing. In regard to optimal amounts of dietary protein, the bulk of the evidence from tissue and animal studies and from human dietary interventions provides a compelling case for the therapeutic effects of high protein diets.
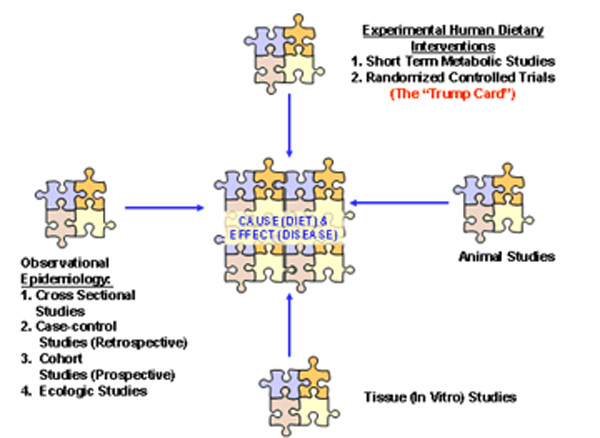
Figure 4. Analytical procedures for establishing cause and effect between diet and disease
Dietary Protein and Cardiovascular Disease
One of the reasons why observational epidemiological studies yield contradictory results is because of the influence of confounding variables which cause confusion in the interpretation of the results because of the mixing of effects from two or more variables (68). For example, although some observational studies have shown a positive association between animal protein and cardiovascular disease (CVD), it is entirely possible that this association is spurious because the measurement of animal protein is confounded by another variable that is also linked to CVD. Meat is a major source of animal protein in the U.S. diet (20), but it is also a major source of saturated fat (73). Because meat comes as an inseparable package of (protein + saturated fat), animal protein ingestion will be highly correlated to saturated fat, thereby making it difficult to disengage the atherogenic effect of saturated fat from that of animal protein. Accordingly, experimental studies are more useful to determine the true effect animal protein may have upon cardiovascular risk factors because they can be designed to isolate the protein effects from the saturated fat effects.
Sinclair and colleagues (74) performed an experiment in which they fed 10 adults a low fat, lean beef-based diet for 5 weeks. Energy intake was kept constant over the 5 week study. Total blood cholesterol concentrations fell significantly within 1 week of commencing the diet, but rose as beef fat drippings were added in a stepwise manner in weeks 4 and 5. The authors concluded, “. . . it is the beef fat, not lean beef itself, that is associated with elevations in cholesterol concentrations”.
Numerous short term human dietary interventions have demonstrated the therapeutic effect of lean, animal based protein upon blood lipid parameters. Wolfe and colleagues have shown that the isocaloric substitution of protein (23% energy) for carbohydrate in moderately hypercholesterolemic subjects resulted in significant decreases in total, LDL and VLDL cholesterol, and triglycerides while HDL cholesterol increased (75). Similar blood lipid changes have been observed in normal healthy subjects (76) and in type II diabetic patients in conjunction with improvements in glucose and insulin metabolism (77, 78). A litany of more recent studies has confirmed that elevations in dietary protein have a beneficial effect upon blood lipid profiles (79-85). The mechanism or mechanisms of action of high protein diets upon blood lipid chemistry are not clear; however animal studies suggest that the beneficial effects are caused by their powerful inhibition of hepatic VLDL synthesis, perhaps by altering apoprotein synthesis and assembly in the liver (86).
The relationship between protein intake and blood pressure has been comprehensively examined in observational population studies, which support the notion that higher protein intake can lower blood pressure (87-89). A substantial number of randomized controlled trials have demonstrated that higher dietary protein either from soy (90-92), mixed dietary sources (85) or from lean red meat (93) significantly lower blood pressure.
Dietary Protein and Insulin/Glucose Metabolism and Weight Regulation
In addition to reducing CVD risk by improving the blood lipid profile and reducing blood pressure, higher protein diets have been shown to improve insulin sensitivity and glycemic control (79, 81, 84, 94-96 ) while promoting greater weight loss (80, 83, 84, 97, 98) and improved long term sustained weight maintenance (99, 100) than low fat high carbohydrate calorie restricted diets. The weight loss superiority of higher protein, calorie restricted diets over either calorie restricted (low fat/ high carbohydrate) diets or calorie restricted (high fat/low carbohydrate) appears to be caused by the greater satiety value of protein compared to either fat or carbohydrate (97, 100-103). Of the three macronutrients (protein, fat, carbohydrate), protein causes the greatest release of a gut hormone (PYY) that reduces hunger (103) while simultaneously improving central nervous system sensitivity to leptin (97), another hormone that controls appetite and body weight regulation.
Dietary Protein and Bone Health
One of the crucial issues regulating bone mineral health and integrity is calcium balance which represents the difference between the amount of dietary calcium which is absorbed and the amount of calcium leaving the body through the urine and feces. Figure 5 demonstrates two key points: 1) most (~75%) of dietary calcium is not absorbed, and 2) calcium absorption increases with decreasing dietary intakes and decreases with increasing dietary intakes (104). Because dietary protein has been frequently, but not always (105-108), shown to increase urinary calcium excretion, it is possible that long term ingestion of high protein diets could lead to accelerated loss of calcium from the bones thereby impairing bone health and integrity.
Without the concurrent measurement of dietary calcium absorption along with urinary calcium losses the net calcium balance cannot be known. Hence, the simple observation that dietary protein ingestion may increase urinary calcium losses tells us little or nothing about calcium balance. In evaluating the effect of high protein diets upon bone mineral health, it is therefore crucial to measure both urinary calcium excretion and intestinal absorption of calcium. In this regard, Pannemans and colleagues (109) compared a low protein (12 % energy) to a high protein diet (21 % energy) in young and elderly subjects. Both a higher urinary calcium excretion and a higher intestinal absorption of calcium were induced by the high protein diet, thus no negative calcium balance occurred. A similar experiment confirmed that elevated dietary protein enhances calcium absorption and thereby counters the increased urinary excretion of calcium (110). Furthermore, a series of recent dietary interventions in humans has shown that high protein, meat based diets do not cause loss of calcium from the skeleton, but actually have a favorable effect upon it by lowering bone resorption (105, 107, 111, 112) and may actually increase bone formation by dietary protein induced increases in IGF-1 (105).
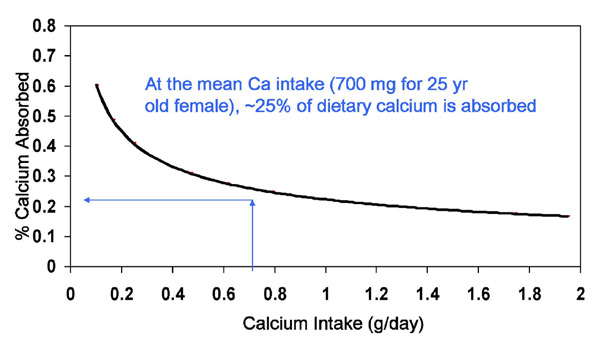
Figure 5. Relation between Calcium Intake and Absorption
Dietary Protein and Kidney Function
One of the most common misperceptions about high protein diets is that they can damage the kidneys of healthy normal individuals. This concept is known as the “Brenner Hypothesis” (113) and suggests that increased dietary protein elevates the kidney’s filtration rate (GFR) which in turn alters the kidney’s structure (glomerulosclerosis) which then causes albumin to appear in the urine (microalbuminuria). Although these series of steps represent the hypothesis Brenner proffered (113), his experiments actually showed an entirely different series of events. In reality, Brenner demonstrated that patients with pre-existing kidney disease had an elevated GFR, glomerulosclerosis and microalbuminuria and that by reducing dietary protein the GFR and microalbuminuria could be lowered (113). He further suggested that because elevated dietary protein increased the GFR in short term studies (< 2 weeks) of healthy normal subjects, protein was responsible for kidney damage. The problem with this interpretation is that markers of functional kidney damage in the normal subjects (microalbuminuria) were not demonstrated along with the elevations in GFR, nor were any long term studies (3-6 months) carried out to determine if the kidneys adapted to a higher protein intake.
The incidence of diabetic end stage kidney disease has increased steadily over the past three decades (114, 115). If dietary protein were responsible for causing kidney damage, then one might expect that dietary protein would have steadily increased during this same time interval. In fact, dietary protein significantly declined during this same period (19). Further, in the National Health and Nutrition Education Survey (NHANES III) of more than 12,000 subjects, dietary protein intake was unrelated to the incidence of microalbuminuria in normal, healthy subjects (117). In a comprehensive review of all randomized controlled trials evaluating the effectiveness of low protein diets upon GFR and kidney function, 11 studies (total n = 1223) showed no therapeutic effect, while only three investigations (total n = 235) demonstrated a significant benefit of dietary protein restriction on kidney failure progression (117).
The reason high protein diets do not have adverse effects upon kidney function in healthy, normal subjects may lie in the ability of the kidney to adapt to a higher protein intake. In a randomized controlled dietary intervention lasting 6 months, Skov and colleagues evaluated the effect of high (25 % energy) and low (12 % energy) protein diets upon kidney function in 65 healthy male and female subjects (118). The high protein diet did not cause urinary albumin to increase. Additionally, the specific GFR, which is an expression of the filtration rate per unit kidney volume, did not change during the high protein diet, indicating that renal (kidney) adaptation occurred to the higher protein load. The authors summarized, “We therefore conclude that a high dietary protein intake does not appear to have adverse effects on renal function in individuals without renal impairment.”
Dietary Protein and Cancer
Observational epidemiological studies frequently (119), but not always (120) show that high animal protein diets may increase the risk for a variety of cancers, particularly colorectal cancer (121). Consequently, it might be expected that non-meat eating vegetarians would have a lower risk for these cancers. Paradoxically, this effect has not been consistently demonstrated (119). A proposed mechanism of action for the carcinogenic effect of meat consumption is the formation of toxic N-nitroso compounds (NOC) in the gut from heme iron in meat (122, 123). Short term human studies are in agreement that increased meat consumption increases NOC formation both in the lower (122) and upper (123) gastrointestinal tract. However, whether this situation translates into increased cancer risk is not known because to date, no randomized controlled trials of increased meat consumption in humans, using cancer diagnosis as an end point, have been conducted.
The meats and fish consumed by pre-agricultural humans were almost always fresh, whereas current western diets contain significant quantities of processed, salted meats and fish preserved with nitrites and nitrates. Processed meats contains 10 times more NOC (5.5 µmol/kg) than fresh meat (0.5 µmol/kg) (124). Pre-agricultural humans consumed their fresh meats along with high intakes of fresh fruits and vegetables estimated to be between 35-45 % of total energy (14) compared to 8.1 % of total energy in the current U.S. diet (125). Increased fruit and vegetable consumption increases the fecal transit rate so that NOC have less contact time with the colonic mucosa and therefore may reduce the carcinogenic risk (126). Hence, the context under which high meat consumption occurred in hunter-gatherers varied significantly from what occurs in westernized populations. Animal based foods were almost always consumed fresh in conjunction with copious quantities of fresh fruits and vegetables. Even when vegetable intake was low or absent in these peoples, there is little evidence for an association of high protein, animal based diets with colorectal cancer. Prior to western acculturation, the Inuit may have consumed more than 95 % of their daily energy from animal and seafood (15), yet a comprehensive review examining virtually all historical and ethnographic data of these people prior to westernization was unable to document a single case of colorectal cancer (126). Should a high protein meat based diet initiate or promote colorectal cancer, then one might expect obligate carnivores such as cats to demonstrate high incidences of these malignancies. In, fact the opposite is true, and the rate of gastrointestinal tract cancers is quite low in domestic cats (128). In summary the case for animal based, high protein diets causing colorectal cancer, within the context of pre-agricultural diets, is weak.
Dietary Protein and Muscle Protein Synthesis and Fatigue
For athletes and individuals engaging in regular exercise, an animal based, high protein diet may be ergogenic and facilitate improved performance because of the stimulatory effect of dietary branch chain amino acids (BCCA) upon muscle protein synthesis (129-131), particularly when they are consumed in the post exercise window (132, 133). Table 2 demonstrates that lean meats and fish are much richer sources of the branch chain amino acids (valine, leucine and isoleucine) than are plant foods. In addition to facilitating muscle synthesis during the post exercise recovery period, BCCA may also improve endurance performance by reducing perceived exertion and mental fatigue by reducing the synthesis of brain 5-hydroxytryptamine, a substance that may promote central fatigue (134).
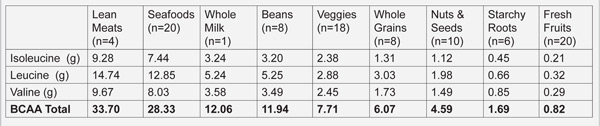
Table 2. Average branch chain amino acid (isoleucine, leucine and valine) concentration in various food groups (1000 kcal samples)
Dietary Protein: Summary and Conclusions
The evolutionary evidence indicates that so called “high protein diets” (20 – 30 % total energy) and “very high protein diets” (30- 40 % total energy) actually represent the norm which conditioned the present day human genome over more than 2 million years of evolutionary experience. The evolutionary template would predict that human health and well being will suffer when dietary intakes fall outside this range. Hence the current U.S. consumption of protein (15 % total energy) may not optimally promote health and well being. There is now a large body of experimental evidence increasingly demonstrating that a higher intake of lean animal protein reduces the risk for cardiovascular disease, hypertension, dyslipidemia, obesity, insulin resistance, and osteoporosis while not impairing kidney function.
How Much Protein Is Needed?
T. Colin Campbell, PhD
Jacob Gould Schurman Professor Emeritus Of Nutritional Biochemistry
Cornell University
Determining how much protein to consume is not a simple exercise. Protein, which is distinguished from its macronutrient carbohydrate and lipid partners by its nitrogen (N) content, is an essential nutrient. Philosophical, cultural and economic concerns have influenced the discussion on how much protein to consume ever since it was first discovered and named in 1839 by the Dutch chemist, Gerhard Mulder.
A good place to begin understanding how much protein to consume is with the recommended daily allowance (RDA, now recommended daily intake, RDI) that has been repeatedly re-evaluated and re-established for the past 60-plus years by many official bodies. Probably the simplest method of determining recommended intakes, at least conceptually, has been the N balance study. An experimental determination is made of how much protein, analyzed according to its N content, must be consumed in order to compensate for the average daily amount of N lost from the body.
After adjusting for the 16% N content of protein and after adjusting upward the experimental mean by two standard deviations to include the needs of about 98% of the larger population, this gives an ‘allowance’, or recommendation, of 0.8 gm of protein to be consumed per day per kg of body weight, for both sexes. This is equivalent to 48 gm of protein for a 60 kg adult female (132 lb), or 56 gm for a 70 kg (154 lb) adult male. Because this is based on the same dietary protein level for both sexes, I prefer these amounts to be expressed as percent of total diet calories. Along with the consumption of 0.8 gm of protein per kg/d, about 35 calories per kg/d is also consumed (i.e., 2450 calories for a 70 kg person). Therefore, 56 gm of protein per day represents 224 calories (1 gm of protein = 4 calories), or only 9.1% of the total 2450 calories consumed per day.
Said another way, a 9% protein diet theoretically is in excess of the amount of protein needed by about 98% of the population. Although some adjustments have been made for growth, pregnancy and lactation, a dietary protein content of about 10% is generally considered adequate for good health, for both sexes and for all ages.
The question, then, should not be how much protein is needed but how much protein in excess of the 10% dietary level can be safely consumed. This a highly relevant question because, according to national surveys, diets containing from 11% to 22% protein (mean of about 16%) are routinely consumed by about 90-95% of the American population. Said another way, the question is what health benefits and/or risks might be expected by consuming protein in excess of the amount needed?
At this point, it is worth noting that most people, for a very long time, have been concerned about getting enough protein. Many have also assumed that animal-based foods are the best source of protein because of their higher ‘quality’ or ‘biological value’, an underlying assumption of my own doctoral research dissertation about 45-50 years ago. The scientific community has indeed encouraged this public assumption ever since the discovery of protein. Mulder, for example, in 1839 gave this N-containing substance its name from the Greek word, proteios, which means ‘of prime importance’. Justis von Liebig, a contemporary of Mulder, went on to state that protein was the stuff of “life itself”. The famous German nutritional chemist Karl Voit and his many prominent students in the late 1800s and early 1900s, recommended that 100-130 gm per day should be consumed even though Voit, himself, found that 52 gm per day was sufficient for good health. Voit’s student, Max Rubner, also a prominent nutrition researcher, said that protein interchange was “the right of civilized man” while another Voit student, A.O. Atwater, went on to encourage similarly high protein intakes while he was founding the USDA laboratory in the U.S.
Importantly, these early nutrition scientists, ultimately becoming ‘fathers’ of their field, almost always equated protein with animal flesh. Although they acknowledged that plants also had protein, it was of “lower” quality, so they said, thus laying the groundwork for the idea that plant proteins had nutritional value only when they were carefully combined in order to compensate for their respective deficiencies of certain amino acids.
The personal biases of these early investigators often were expressed with considerable hostility and arrogance. The medical director of the British Empire Office in India, for example, offered that those who live on plant-based foods were of a “cringing and effeminate nature”.
In sharp contrast, Professor Russell Chittenden of Yale University School of Medicine, offered a very different set of findings, as published in two large books in 1904 and 1907. He showed that a group of young military recruits could obtain excellent physical conditioning over a 6 month period, as examined in 15 strength and endurance tests, by consuming a mostly plant protein-based diet. Not to be misunderstood that it might be the training and not the diet that led to these results, he then showed even greater conditioning when a group of already fit athletes switched to a plant-based diet. These remarkable and highly significant findings came at a time when Voit and his students and associates were pushing for higher, not lower, intakes of animal-based protein. Later, in 1922, the famed Dr. Benjamin Spock who was at that time a member of the Olympic crew team at Yale, was being advised along with his team mates to consume a high animal protein diet. He learned of Professor Chittenden’s work at his university only many years later when he wrote to me shortly before his passing and wondered why his coaches had made no mention of their colleague’s work. Little did Spock know that, about that same time in the early 1920s, Chittenden was being severely chastised for his work by his professional colleagues. Was he, perhaps, of an effeminate nature?
Let’s now return to the question of what happens when protein intakes are increased beyond 10% of calories, say from about 10% to 20% of calories, the range for most Americans. A variety of adverse health effects have been demonstrated and these effects are remarkably consistent among clinical, experimental (i.e., intervention), and observational type studies.
At the clinical level, for example, animal-based proteins (primarily) act on fundamental biological systems such as acid-base buffering, hormonal messaging, and enzyme activities that set the stage for a wide variety of adverse health outcomes. Thousands of studies have demonstrated these effects.
For example, the consumption of animal-based proteins can lead to a chronic condition known as metabolic acidosis, which has been known since the late 1800s. Secondary effects of this condition are extensive and include, for example, bone calcium loss to buffer the excess acid (thus weakening the bones) and inhibition of the critical enzyme that controls the formation of the ultimate vitamin D metabolite now known to be involved in a wide variety of health promoting effects.
Hormonal profile distortion, for example, includes elevated estrogenic hormone activities (higher estradiol and lower sex hormone binding globulin) and its corollary effects on breast cancer risk and other steroid responsive tissues. Also, animal protein consumption increases growth hormone activities like IGF-1 and IGF-2 that now associate with the promotion of multiple cancers.
Altered enzyme activities can be quite dramatic, occurring within hours after consumption of increased amounts of animal-based proteins and, further, these effects can be sustained for long periods of time. These dietary protein effects are especially notable for an enzyme complex involved in a broad array of reactions, including chemical carcinogen activation, steroid biosynthesis and disposition and fatty acid metabolism.
These clinical effects are consistent both with human intervention studies designed to control or even reverse serious chronic diseases and with human population studies. Increasing intakes of animal protein within the 10% to 20% range increases urinary calcium loss, elevates total and LDL cholesterol levels, increases cell replication and enhances DNA damage resulting from enzymatic activation of chemical carcinogens. These and many other clinical effects parallel higher incidence and/or mortality rates of osteoporosis, cardiovascular diseases, certain autoimmune diseases and various cancers, among other ailments. These findings are only a small sample of animal protein induced disorders and are cited here because of the consistency of evidence across clinical, experimental and observation type studies within the 10-20% range of dietary protein experienced by most Americans.
Although these outcomes and observations mostly refer to the independent effects of protein, it should be noted that the isolated effects of individual nutrients usually represent only a small part of a much larger story. First, the activities of nutrients studied in isolation are often quite different than when they are studied in the context of food. Second, when animal protein-based food consumption is increased, plant protein-based food consumption is usually decreased. Thus associations of animal-based foods with various disease outcomes almost invariably include simultaneous contributions resulting from the absence of the protective factors in plant-based foods. Plant-based foods have a wide variety of constituents that tend to oppose, or re-balance, the adverse effects of animal-based foods on these fundamentally important biological systems.
These are only a few of the effects of increasing total dietary protein above 10% of diet calories, mostly by the addition of animal protein-based foods. But these findings are cited here because (1) they show effects of animal-based protein on fundamental biological systems, (2) they are consistent across different types of studies and (3) they occur within the 10-20% dietary protein range. What makes this more generalized information especially compelling is its ability to explain a group of diverse yet remarkably profound diet and health observations reported during the past few decades. In population studies, for example, animal protein intake exhibits very impressive correlations (80-90%) with breast, prostate, ovarian, kidney and colon cancer, with increased disease risks appearing at very low levels of dietary animal protein (i.e., the regression line passes through the origin). Similar correlations also exist with cardiovascular diseases, type I and II diabetes, hypertension, osteoporosis, multiple sclerosis and kidney stones, among other ailments—all within the 10-20% dietary protein range.
Moreover, dramatic reversal, even cure, of advanced degenerative diseases as well as relatively non-fatal ailments, have been observed by switching away from animal protein-based foods to whole plant protein-based foods. The demonstrated reversal of advanced heart disease in humans is truly astounding, as is the 34 years-long control of multiple sclerosis progression. And finally, our own years-long series of experiments with laboratory rodents are especially informative of the fundamental effects of animal protein on experimental cancer development. When casein (cow’s milk protein), for example, is fed above the level that meets physiological needs, tumor growth progresses; when casein is fed at levels below 10%, tumor growth is suspended or reversed. That is, we could turn on and turn off experimental tumor development by feeding or withdrawing dietary casein at levels above or below 10% of calories, respectively.
I am convinced that many of the most cherished assumptions of nutritional science can trace their origin to our long and unquestioned reverence for protein, especially for animal-based protein. Diets have been constructed and judged according to their inclusion of this treasured nutrient. Diet and health policy has been structured in such a way so as to protect the consumption of animal-based protein. The first ‘official’ recommendation by the National Academy of Sciences in 1982 to decrease fat intake only to 30% of diet calories—even though the evidence at that time indicated lower fat intake was warranted—was to protect the consumption of animal protein-based foods. I know because, as a member of that committee, I wrote with a colleague the draft chapter on protein and cancer showing that animal protein intake was as much associated with cancer risk as was fat intake.
This long-standing reverence for animal-based protein often seems to be very personal, perhaps macho building, for some individuals. In point of fact, it may surprise many that when done right, consuming whole plant-based foods are known to lead to better not lesser athletic performance and physical fitness. Nonetheless, questioning the health value of animal-based protein still causes as much hostility today as before for some individuals, especially if they are economically or religiously conflicted.
My conclusion from these many observations is that animal protein, when added to diets already containing 10% protein, has the potential to promote the development of a wide variety of serious and oftentimes fatal diseases—not only because of the direct effects of protein but also because of the many parallel effects of companion nutrient imbalances created by animal-based foods simply to get access to the extra protein. I find that 10% total dietary protein, which has been long recommended by multiple policy committees and acknowledged in scientific reviews, is not only adequate but also is relatively devoid of risk of serious diseases and other ailments. It just so happens that 10% dietary protein is the same level that is typically found in a diet of varied whole plant-based foods. Increasing dietary protein above this 10% level generally means adding or substituting animal-based foods for plant-based foods but doing so invites a plethora of adverse health effects. To be very clear, I believe that total dietary protein should be 10% of calories, with virtually all of it being in the form of plant-based protein.
This commentary primarily addresses the effects of dietary animal protein because this was the mandate given to me. But I should emphasize that among the various adverse dietary effects associated with increasing consumption of animal protein-based foods, there is often the possibility that these effects are attributed, at least in part, to the consumption of highly processed foods rich in fat, salt and refined carbohydrates. In many studies, it is not easy to differentiate these separate effects, in part because of inadequate data. Nonetheless, prominent animal protein effects have been observed and mechanisms have been defined for the singular effects of this nutrient. In the final analysis, however, consuming a diet comprised of whole plant-based foods solves both problems caused by animal-based and highly processed foods. It is clear to me that a diet comprised of whole vegetables, fruits and cereal grains (unless otherwise contraindicated by allergic responses) with little or no animal-based foods, resulting in a dietary protein level of about 10%, is as near to an ideal diet as one can get.
Rebuttal to:
T. Colin Campbell, Ph.D.: How Much Protein Is Needed?
Loren Cordain, Ph.D., Professor
Department of Health and Exercise Science
Colorado State University
Rebuttal Overview
For the readers I think it is necessary to state that I have no personal enmity towards Colin – rather the opposite. I have met him at a nutritional conference in Boulder, Colorado a few years ago, and we had a warm and cordial conversation. My sole goal with my first paper and with this rebuttal is to seek the most accurate scientific truth that the available data can provide regarding the role dietary protein plays upon human health. Above all, the data must speak for itself without the intervening prejudice introduced by charismatic personalities, faulty human judgment and preconceived biases. I repeat: the data must speak for itself. Scientific theories must solely be driven by informed hypotheses based upon objective, well controlled experimental data. When the data consistently do not support the hypothesis, it is inappropriate to manipulate and selectively use flawed data (either knowingly or unknowingly) to continue to support the erroneous hypothesis (135). Initially, no matter how contrary they may appear to common sense, new hypotheses must be tested to replace previous hypotheses that no longer are consistent with the best available data (135).
I have organized my rebuttal into two categories: 1) general comments, and 2) specific comments. Within the general comments I will point out the generally flawed scientific logic that Colin follows to form his hypothesis that high protein diets have deleterious health effects. Within the specific comments, I will show you, point by point, how Colin has overstepped the bounds of the scientific method by misinterpreting data and selectively ignoring key studies that demonstrate the therapeutic effect of high protein diets.
In this rebuttal I have employed the reference numbering system I used in my opening paper, and any additional references I use in this rebuttal begin with the number, 135.
General Comments
The scientific study of human nutrition represents a sub-discipline within the much greater discipline of biology. Over the past 100 years, remarkable “discoveries” have been made into how dietary factors influence our health and well being. For instance, the “discovery” that vitamin B12 was an essential component of the human diet, without which our health suffers greatly, was made as recently as 1948 (1). The “discovery” that diets devoid of animal food cause blood and urine concentrations of the conditionally essential amino acid, taurine, to decline has only been known since 1988 (40).
A more accurate description of “discoveries” in biological sciences (in which the study of nutrition falls) is the uncovering of pre-existing phenomenon. The human requirement for vitamin B12 has been in place for eons before we uncovered what was already there. The human requirement for vitamin B12 and the decline in blood concentrations of taurine with reduced animal protein occur, not because scientists have “discovered” them, but rather because these fundamental mechanisms are woven into the fabric of our genes. Humans have a dietary requirement for vitamin C because we lack a functional gene that codes for the enzyme (L-Gulono- γ- lactone oxidase) which normally allows glucose to be converted to vitamin C. Analysis of the human gene coding for L-Gulono- γ- lactone oxidase shows that it has acquired a number of mutations (via natural selection) over the past 70 million years which have rendered it inactive (136). Because our primate ancestors ingested sufficient vitamin C containing foods, mutations which inactivated this gene had little negative influence upon survival and reproductive success of individuals bearing these genes.
Any unified theory of human nutrition represents a detective story in which scientists attempt to reveal or uncover physiological systems that have been designed by, and put in place by evolution through natural selection. Accordingly, hypotheses regarding what modern day humans should and shouldn’t eat must be consistent with the system and the ancient environmental selective pressures that engineered our current genes. If we are to buy into Colin’s hypothesis that optimal human health occurs when dietary protein represents about 10% of energy, then the system (evolution via natural selection) which shaped our present genome necessarily had to be conditioned over eons by a low protein intake. Otherwise, there is no rational alternative hypothesis to explain why humans have a low dietary protein requirement. A low dietary protein requirement for optimal human health can only occur if it is dictated by our present day genes which in turn are shaped by events that have occurred in the environment of our ancestors in the distant and remote past.
As I have laboriously and meticulously lain out in my initial essay, there is no credible fossil, archeological, anthropological, anatomical, ethnographic or biochemical evidence to show that members of our genus (Homo) routinely consumed low protein diets. In fact, without the inclusion of energetically dense animal food into the hominin diet, starting at least 2.5 million years ago, our large energetically active brains would not have evolved (16, 34, 35). Accordingly, the fundamental logic underlying Colin’s hypothesis (that low protein diets improve human health) is untenable and inconsistent with the evolution of our own species.
Specific Comments
1. Page 18, column 1, paragraph 1. As Colin has suggested, nitrogen balance studies have been extensively used to predict the minimal human protein requirements (137). Because all protein contains 16.6% nitrogen, then the measurement of nitrogen serves as a proxy for protein. Colin’s argument is that optimal health occurs when we are in zero nitrogen balance (e.g. when our daily nitrogen intake = daily nitrogen excretion), and that excessive protein intake and hence negative nitrogen balance (nitrogen intake < nitrogen excretion) causes ill health and disease. Colin suggests that zero nitrogen balance occurs when the daily protein intake is 10 % of energy.
The problem with this oversimplification is that minimal protein requirements provide no information about optimal protein requirements. As is the case with many nutrients, the minimal protein requirement may not necessarily represent the optimal requirement. The minimal vitamin D requirement is 600 IU, however additional therapeutic effects occur when the daily intake is considerably greater (138, 139). Similarly, niacin (vitamin B3) is of therapeutic benefit in lowering blood cholesterol concentrations at daily intakes much higher than daily recommended intakes (DRI) (140). As I have extensively outlined in my first paper, a large body of experimental evidence now demonstrates a higher intake of lean animal protein reduces the risk for gout (49), cardiovascular disease (70, 74-86), hypertension (85, 87-93), dyslipidemia (74-86), obesity (80, 83, 84, 97-103), insulin resistance (79, 81, 84, 94-96 ), and osteoporosis (105, 107, 109-112 ) while not impairing kidney function (116-118).
2. Page 18, column 2, paragraph 4. It is difficult to objectively analyze experiments conducted more than 100 years ago in regard to their present day validity (141). As Colin has indicated, the design of this experiment was such that the exercise training effect could not be separated from the dietary effect. Hence the improvement in strength and endurance shown in this experiment cannot necessarily be attributed to the plant based diet, particularly when more recent, well controlled studies have shown otherwise.
As I have previously pointed out in my original essay, plant based diets contain much lower concentrations of the ergogenic branch chain amino acids (BCAA) (valine, leucine and isoleucine). For instance, a 1000 kcal serving of lean meat contains on average 33.7 grams of BCAA whereas a comparable serving of whole grains contain a paltry 6.1 grams of BCAA. High protein diets are ergogenic because of the stimulatory effect of dietary BCCA upon muscle protein synthesis (129-131), particularly when they are consumed in the post exercise window (132, 133). Additionally, BCCA may also improve endurance performance by reducing perceived exertion and mental fatigue by reducing the synthesis of brain 5-hydroxytryptamine, a substance that may promote central fatigue (134).
3. Page 19, column 1, paragraph 3. In regard to the metabolic acidosis produced by meats and their potential for adverse health effects, it should also be pointed out that cereal grains (a foundation of plant based diets) maintain an acid load similar to meat and animal foods (142). Whereas lean animal protein consumption has been shown to have a favorable effect upon bone mineral health by lowering bone resorption (105, 107, 111, 112) and increasing formation (105), whole grain cereal consumption impairs bone mineral health (143). In populations where cereal grains provide the major source of calories, osteomalacia, rickets and osteoporosis are common place (144-147). In animal studies it has been long recognized that excessive consumption of cereal grains can induce vitamin D deficiencies and bone mineral abnormalities in a wide variety of animals (148-150) including primates (151). The mechanism by which cereal grain consumption promotes bone mineral disorders may be via their ability to interfere with the entero-hepatic circulation of vitamin D or its metabolites (151, 152), or by increasing the rate of inactivation of vitamin D in the liver (153).
4. Page 19, column 1, paragraph 6. As I have previously pointed out, increased loss of urinary calcium by high protein animal based diets does not necessarily result in skeletal calcium loss (105, 107, 111, 112 ) because calcium absorption is simultaneously increased (109, 110). In regard to lean animal protein elevating total and LDL cholesterol, I am not aware of a single study which has demonstrated this effect, particularly when the effect of saturated fat has been eliminated. In fact, the opposite is true. Increasingly, experimental evidence in humans shows that lean animal protein improves all facets of the blood lipid profile (79-85 ).
5. Page 19, column 2, paragraph 2. As I have previously pointed out in my initial essay, observational epidemiological studies cannot show causality (68, 69). In addition, not all epidemiological studies show strong or “impressive” relationships between animal protein consumption and various cancers (120, 154, 155), particularly when the effect of high cooking temperatures is eliminated (156). Observational epidemiological studies showing “correlations” to type 2 diabetes, hypertension, osteoporosis and kidney stones fall by the wayside and become meaningless when more powerful human dietary interventions show that high protein diets are actually protective for these diseases and their symptoms, as I have extensively reported in my first essay.
6. Page 19, column 2, paragraph 3. In this paragraph Colin suggests that reversal of a number of degenerative diseases can occur when animal protein based foods are replaced with whole plant protein based foods. Unfortunately, no references are provided so it is difficult to comment or rebut these remarks. However, I can comment upon the extensive rodent experiments that were conducted in Colin’s laboratory over more than a decade in which he examined the role of milk protein (casein) in the development of liver cancer.
Aflatoxins are naturally occurring toxins that are produced by a variety of fungi, most notably Aspergillus flavus and Aspergillus parasiticus. These fungi normally reside in soil, but can invade food crops under conditions of high humidity or other stress. Food crops most frequently affected are cereals, nuts and legumes (which represent the mainstays of plant based, vegetarian diets). The toxin is also found in the milk of animals fed contaminated feed. Aflatoxins are metabolized in the liver to become potent liver carcinogens for all mammals including humans (157). High-level aflatoxin exposure causes liver cancer, whereas chronic, low level exposure to aflatoxin does not necessarily lead to cancer (157).
Colin’s research group developed a rodent model of liver cancer in which they dosed the animals with high concentrations (typically 200-350 microgram/kg per day for 10 days) of aflatoxin and then fed them diets containing varying amounts (typically 20, 16, 12, 8, 4 % of total energy) of casein (158-161). Regardless of the casein dose, all animals developed cancerous or pre-cancerous liver lesions (161), however the animals fed the higher amounts of casein developed more cancerous lesions, particularly when a level of approximately 12 % casein was reached (160). From these experiments Colin concluded that, “low protein intake inhibits lesion development” and that “AFB1 (aflatoxin) induced preneoplastic foci (precancerous lesions) depends upon a high protein intake” (161).
Although Colin has inferred from his experiments with rodents that high protein diets promote cancer and low protein diets repress it following cancer initiation by a carcinogen, this interpretation is incorrect. The only logical conclusion that can be reached from his series of experiments is that only the milk protein, casein, when consumed at more than 10% of energy, promotes liver cancer in rodents exposed to high concentrations of aflatoxin. His experiments cannot be generalized to other animal proteins, such as those found in lean meats. In the typical U.S. diet, dairy food consumption represents 10.6 % of the total energy (13) and is distributed in the following manner: whole milk (1.6 %), low fat milk (2.1%), cheese (3.2 %), butter (1.1 %), and other 2.6 % (13). The total protein content by energy for milk is 21 % , low fat milk ~29 % , cheese ~25 % , butter 0 %, and other ~27 %. Casein represents ~80 % of all milk proteins, so the average total casein content in the U.S diet is ~2.0 % energy. Accordingly, current consumption of casein in the U.S. diet would have little or no bearing on cancer incidence rates if we assume Colin’s rodent model of cancer is correct and applicable to humans.
In the typical U.S. diet ~75 % of the protein comes from animal products other than dairy (20). In the U.S. diet muscle meats from birds, mammals, fish and invertebrates represent far and away the greatest protein source, as organ meats are infrequently consumed. The primary proteins in muscle tissues are actin and myosin. Consequently, Colin’s rodent experiments using casein as a generalized surrogate for protein in the typical U.S. diet has little or no relevance to human cancers for two reasons. First, as I have previously shown, in the U.S. population it would be virtually impossible to ingest 10 % of the daily caloric intake as casein, and secondly the primary animal proteins (actin and myosin) in the typical U.S. diet were never tested in Colin’s animal experiments. Hence Colin’s conclusions that high dietary protein from all animal sources promotes cancer and lower dietary protein prevents it cannot be inferred from his rodent experiments.
7. Page 20, column 2, paragraph. Colin’s statement “It is clear to me that a diet comprised of whole vegetables, fruits and cereal grains (unless otherwise contraindicated by allergic responses) with little or no animal based foods, resulting in a dietary protein level of about 10%, is as near to an ideal diet as one can get” clearly reflects the bias he brings into this debate. If only Colin would have framed his hypothesis from an evolutionary perspective years ago, he would have realized that his viewpoint of an ideal diet is erroneous and untenable.
It may surprise Colin, but cereal grains were rarely or never consumed by all of humanity until very recent times (< 500 human generations) (13, 143). Additionally, the incidence of celiac disease (caused by consumption of wheat) is now estimated to afflict 1 in 133 people in the U.S. (162). What should the ideal diet be for these 2,255,639 U.S citizens? And why might autoimmune diseases be caused by grain consumption in the first place (143, 163)? Finally, if vegetarian diets are “as near to an ideal diet as one can get” why then do meta-analyses of all cause mortality (including cancers) in vegetarians show them to be no better off than the general public? (164).
Rebuttal to:
Loren Cordain, PhD: The Evolutionary Basis for the Therapeutic Effects of High Protein Diets
T. Colin Campbell, PhD
Jacob Gould Schurman Professor Emeritus of Nutritional Biochemistry
Cornell University
My critique of Professor Loren Cordain’s proposition almost entirely depends on my philosophy of nutrition. It is clearly different from that of Cordain’s understanding of this discipline. He mocks the science of nutrition as if it has little or nothing to offer. I believe that it has much to offer even though its essence mostly remains hidden. He says that nutritional science is a “newly established discipline”, that it “is a highly fractionated, contentious field with constantly changing viewpoints”, that it “remains an immature science” and “that there is such seeming chaos, disagreement and confusion”. He then suggests “nothing in nutrition seems to make sense because most nutritionists have little or no formal training in evolutionary theory”. He apparently believes that only paleontologists and archeologists have a crystal ball as to what is nutrition and, more specifically, how much protein should be consumed.
He has thrown down a challenging gauntlet, thus leaving me no option but to explain what I believe nutrition to be—before I tackle the protein requirement question.
While I agree that there is enormous confusion, both in the professional and lay communities, I disagree that nutrition is a relatively “new” science and that hypotheses about nutrition must pass a test set by archeologists before they can be considered reliable. I agree that nutrition hypotheses should be consistent with our evolutionary past, but having to pass an archeologist’s ‘smell’ test is too restrictive based on the evidence presented in Cordain’s paper.
Protein was discovered in 1839, fat and energy were being discussed in the mid 1800s and amino acids were known to be components of protein in the late 1800s. I suggest that nutrition as a science predates the well-known disciplines of genetics, bacteriology, virology, biochemistry, pharmacology, immunology and molecular biology, to name only a few. Is it possible that Cordain considers nutrition to be a “new” science because it is new to him? Is it so confusing to him because he knows so little about the science?
Being confused about the concept of nutrition is a serious matter. It is real—both inside and outside of the field. Many factors contribute to this confusion. In addition to explanations like ignorance, economic pressures and food preference biases, I believe that there is a more fundamental issue that underlies each of these explanations, this being the way we think about scientific investigation itself. More specifically, it concerns the role that precision, as a concept, plays in research. Ever since the Renaissance, scientific investigation has emphasized this concept when gathering empirical evidence. Physical measurements must be precise, substances and events must be precisely characterized physically and chemically, and hypotheses have more value if they are precisely focused (especially if one expects to compete for research funding). Even to question this concept of precision is scientific heresy to many scientists.
Making use of precise measurements is essential for the research and development (R&D) of complex physical technologies like automobiles, space shuttles, telephones, and computers. When developing these complex physical products, we precisely make the component parts then precisely assemble them into complex products. We must be mindful of precision and I am delighted for that.
But the same cannot be said for biomedical R&D, even though most researchers and other observers seem to believe otherwise. Although we may precisely know and control the components and activities of a biological event (as with a specific biochemical reaction), this is far, far removed from trying to ‘do’ R&D on complex products that we call life, disease or health. This is also true for ‘simpler’ products like body organs or even sub-cellular organelles.
Of course, there are times when precision in biomedical research is exceptionally important. For example, we can surgically penetrate minute bits of tissue, we can determine the exact chemical structure of each of the 30,000 or so human genes and we can precisely synthesize chemicals (i.e., drugs) that interact with these genes and their protein products. These are fascinating research journeys and sometimes they can produce real life benefits. But no matter how precisely measured and described are these events and their component parts, we cannot assemble them into complex biological outcomes like those fashioned over millions of years by Nature. We cannot construct biological complexity that is faithful to the natural order of things, no matter how precisely we measure and know the component parts of these complex systems. This is because each event or bit of matter of these systems is usually investigated in relative isolation from its native biological environment. In biological systems, concentrations and activities of the component parts do not exist in discreet quantities (as in physical systems) because they operate, in time, over a continuum. And making matters decidedly more complicated, countless higher order interactions among the component parts cannot be accurately described or controlled. When investigating the nature of complex intact biological systems, we cannot construct them de novo as in physical systems. We can only provide resources then simply observe the outcomes that are produced.
In brief, the criteria and considerations for doing research and development on physical systems are very different from doing research on biological systems. With physical systems, we are in control when doing research developing products. Measuring and assembling things with precision is in our best interest. We know the dimensions, the structural identities and the time parameters and we can stabilize them to our specifications. Except in the most narrowly defined systems, we cannot use precision to construct or even to accurately describe the far more complex and dynamic biological systems. Measuring things with precision does not have the same meaning in biology because the system is always changing in exceptionally complex ways. Moreover, the interactions between the component parts are also changing.
By exaggerating the importance of precision in biomedical research, we also err in other ways. That is, we too often misinterpret the concept of precision as perfection. Assuming more perfection implies more ‘value’ and for some scientists, precision may mean invincibility and absolute truth (or might we say ‘arrogance’?). A major impetus for this view of precision, cum perfection cum absolute truth, is the marketplace mentality where health products and information are valued more for their monetary value than for their health value. Monetary value can be measured precisely but, more importantly, intellectual property protection is enhanced with more precise knowledge.
Biomedical and nutritional research should be conducted within a paradigm that is substantially different from the contemporary, traditional paradigm. We must strive to develop hypotheses, to organize research studies, and to interpret and apply results that acknowledge biological complexity and that describe cause-effect relationships cognizant of that complexity. From this perspective, observations on the component parts, however precisely they may be measured, only represent relative truths, not absolute truths. Moreover, we should stress research that is observational, comprehensive and representative of real life conditions.
This same biological complexity describes the essence of nutritional effects or, perhaps, describes what nutritional science should be. Most serious students of nutrition know well this complexity even though many scientists ignore its implications.
The implications of this view are wide-ranging and probably more significant than most people realize. This view is ‘holistic’ and, among other considerations, means that deciding which groups of foods to consume is far more important than deciding what levels of individual nutrients should be consumed. It means that priority should be given to the search for collective disease and health outcomes that may respond to the same diet and lifestyle conditions. It means investigating a variety of metabolic events that mutually support maintaining homeostasis and regulating adaptive responses, then exploring whether there are groups of food to support this group of metabolic events. It means rejecting randomized clinical trials as the acclaimed ‘gold standard’ of diet and health research. It means questioning the validity of recommended intakes/allowances (e.g., RDAs) on a nutrient-by-nutrient basis. It means acknowledging that the functional activity of a nutrient within a food environment can be far greater than an equal chemical amount in a tablet. Most importantly, it means using ‘weight of evidence’—all evidence—as the standard method of determining the reliability of cause-effect hypotheses.
This is my philosophy, both of nutrition and of the scientific method itself. More than any other consideration, this philosophy makes it difficult to comment on a number of Cordain’s assumptions and observations, even if they may be accurately cited. I challenge many of Cordain’s assertions but, in the interest of space, I will comment only on a few of these points in the hope that an explanation of my philosophy will make clear my answers to the others.
Specific Comments
1. Figure 4, page 9. The randomized clinical trial (RCT) is NOT the “trump card” of all diet and health studies, as claimed. RCTs were meant for the testing of pharmaceutical agents and would-be pharmaceuticals called neutriceuticals (i.e., nutrients), not for assessing lifestyle nutrition. RCTs have undoubtedly distorted the concept of nutrition and caused more confusion in diet and health research than almost any other scientific endeavor in recent years.
2. Page 4, second column, last paragraph. The statement that “all human requirements… are ultimately genetically determined” is an oversimplification. Several studies, including a paper of our own over 25 years ago (Science 211:719-721), have shown that in utero exposure to an appropriate agent can cause a non-genetic (i.e., epigenetic) lifetime change that undoubtedly affects nutrient requirements. This finding, now reported in more recent studies by others, would have been incorrectly considered genetic.
3. Page 5, column 1, paragraph 2. The statement is made that “the upper limit of protein ingestion is determined by the liver’s ability to synthesize urea.” If this is an inference that all could be well until this limit is reached, it is grossly misleading. According to Cordain, this equates to an astounding intake of about 246-270 g protein/day, or about 40% of total calories, 4-5 times the long-standing recommendation/allowance! I have 2 problems with this statement.
An adverse nutritional effect almost never results in a single outcome. Such effects occur serially over time, beginning with the least problematic and proceeding to the most problematic. Saturation of the urea disposition cycle is likely very serious but it will be preceded by many earlier adverse effects, some of which may take years to develop. Unfortunately, Cordain glossed over these effects as if they were unimportant. I refer, of course, to the numerous effects that I cited in my paper and that also are present in our book and in my many publications. I am suspecting that he believes that his archeological evidence is sufficient to make his main points and that modern day investigation of existing problems is nutrition gobbly-d-gook.
4. Page 4 and throughout. Cordain laments nutrition researchers as not being schooled in evolutionary theory. Because I was not trained in this discipline, I shall turn for advice to two professionals who were so trained. The first is a creative evolutionary psychologist (PhD, University of Virginia, Instructor at Stanford University), Dr. Doug Lisle, who with his colleague, Dr. Alan Goldhamer, wrote a fascinating small book, “The Pleasure Trap”. As you can see, Lisle also relies on some brilliant work of others (Dawkins and Williams) in this field. Lisle and I have talked about the basis for this high protein recommendation by anthropologists like Loren Cordain and he puts it this way:
“There is an important misconception among those who attempt to use evolutionary theory to guide their thinking in questions about human nutrition: that the diet consumed in human evolutionary history is the diet that optimally serves human health. This is a serious error, and one that makes major mistakes quite possible, even likely.”
Those who follow evolutionary history and who support the inclusion of substantial amounts of animal protein as a recommended dietary practice, make this error. Our dietary evolutionary history, while interesting, absolutely does not yield critical clues for optimal nutritional practices. Human evolution required that our ancestors make dietary choices that maximized gene proliferation. And that is absolutely the ONLY thing that such practices yielded (see Dawkins’, The Extended Phenotype, for clarity on these issues, also G. Williams’ classic Adaptation and Natural Selection).
“The reader may find an example to be helpful. Consider a dietary contest. Suppose that Diet A (a diet nearly exclusively of animal origin) resulted in 100% of individuals living long enough to reproduce, but despite adequate calories available throughout the lifespan, average non-accident/non-violent age at death (thus death was from nutrition-related diseases) was 50 years. Diet B (a diet exclusively of plant origin) resulted in just 60% of individuals living long enough to reproduce (perhaps because they were too finicky to eat meat [or because inadequate calories killed young people]), but if fortunate enough to be presented with an environment of sufficient calories (thus likely avoiding all nutrition related diseases)... life expectancy was 90 years. In this contest, Diet A wins the evolutionary battle, and is adopted. The evidence presented by modern nutritional science suggests that something akin to this may well be the case for humans, who have a large constellation of disease processes that can be laid at the door of animal foods, and effectively reversed with vegan diets (see Ornish, Esselstyn, McDougall, etc.). Seen from this perspective, speculation about human dietary history is a fascinating subject, but is actually irrelevant in the search for optimal nutritional practices. Such practices can only be determined by scientific investigation done in the present time, studying varying dietary practices and determining their comparative outcomes.”
I concur with this view, especially with the inference that present-day conditions need to be studied for present-day dietary advice, including the amount of protein to be consumed. Ample evidence now exists to show that our bodies are continually drawing on diverse non-genetic (i.e., epigenetic) adaptive mechanisms to enhance health and survivability even when the available resources are less than ideal. We adapt to inappropriate levels of nutrient intakes in ways that attenuate the adverse effects otherwise expected. And we do so without assuming that this is accounted for by genetic mutations, as suggested by Cordain (pp. 10-12). This does not, however, mean that we can fully accommodate inappropriate food without harm, only that these adverse effects are less than what they might otherwise be.
Cordain seems to believe that some of the most important evidence favoring high protein diets may be seen by considering the diets of present day hunter-gatherer societies that presumably reflect our evolutionary past. He uses an ethnographic atlas of 229 such societies and finds that “73% of the world’s hunter-gatherers obtained >50 percent of their subsistence from hunted and fished animal foods, whereas only 14 percent of worldwide hunter gatherers obtained >50 percent of their subsistence from gathered plant foods.” These appear to be impressive findings until one reads a rather devastating rebuttal of his findings that was published elsewhere by Professor Katherine Milton at the University of California at Berkeley (Am. J. Clin. Nutr. 71: 665-667). She first points out (1) that, on several grounds, this atlas is not a reliable compilation of dietary information, (2) that, in any event, another author using the same atlas concluded that “gathered vegetable foods were the primary source of subsistence for most of the hunter-gatherer societies he examined”, (3) that some of these “societies are largely free of diseases of civilization regardless of whether a high percentage of dietary energy is supplied by wild animal foods, wild plant foods, or domesticated plant foods taken from a single cultivar”, (4) that non-genetic adaptations are much more important than genetic mutations in determining diet responses, (5) that the “evolutionary history of anthropoid primates” going back 25 million years “shaped their nutrient requirements and digestive physiology well before they were humans or protohumans” (instead of Cordain’s suggestion that it was the last 2.5 million year history that shaped our nutrient requirements), (6) that “there is general agreement that the ancestral line (Hominoidea) giving rise to humans was strongly herbivorous”, being largely based on the consumption of tropical plant foods and (7) that the archeological evidence does not “shed light” on the proportionate consumption levels of animal and plant based foods. These seem to be powerful counter arguments to Cordain’s hypothesis.
Again, our own research supports these views. When metabolic control systems are compared across species as to their responsiveness to food sources and as to their associations with health/disease outcomes, there is overwhelming support for a linkage between the consumption of whole plant based foods, health maintenance, and disease prevention (this diet, not so coincidently, contains about 10% of the total energy as protein). The fact that these systems go deep into our evolutionary past, far more so than even the 25 million year benchmark suggested by Milton, suggests that much of our nutrient requirements could have been fashioned very early in our evolutionary history (certainly more than 2-3 million years ago proposed by Cordain).
Regardless of which side of the issue one favors, I can’t help but wonder whether the discordance Cordain finds in the field of nutrition does not also apply to his own field of evolutionary history. Therefore, why must studies of contemporary diet and health associations by nutrition researchers be submitted for validation by evolutionary history buffs, as suggested by Cordain? Why not the other way round?
5. Page 9, column 1, paragraph 2. I regret to say that Cordain’s commentary on establishing “cause and effect between diet and disease” (p. 18) is markedly different from my views. Aside from my earlier comments on the virtually worthless value for randomized clinical trials for nutrition studies, I also do not agree with his views on cross-sectional, observational studies, as we conducted in rural China.
Traditionally, epidemiologists and others will claim, as does Cordain, that cross-sectional population studies (also called ecological and correlation studies), which compare population characteristics, cannot only be used to generate hypotheses about possible cause-effect variables, not to prove causation. I absolutely agree with this view when one assumes highly reductionist hypotheses. In fact, virtually all epidemiologists and biostatisticians also agree. However, the reductionist philosophy that allows this criticism does not apply in most instances of diet and health associations. In contrast, if one assumes a more holistic philosophy when considering diet-health causality—as one should—then this precaution about the worthiness of cross sectional studies is far too restrictive.
The holistic philosophy may be applied in research studies in at least two ways. First, one can comprehensively record many factors that may influence outcome (even many possible health and disease outcomes) in a search for consistent and biologically plausible causal factors. For example, we can determine the proportion of plant-based food factors (both exposure and metabolic) that are consistently associated in the same study with a particular outcome or group of outcomes. Second, we can interpret the results of such studies in the context of other studies, especially those that show biological plausibility. Many years ago (1991) I explained this view in the 896-page monograph of the China Project, then restated it in our trade book, “The China Study”. It’s unfortunate that Cordain uses old logic to criticize findings like ours without reading the rationale for that study.
6. Page 6, column 1, paragraph 3. I do not subscribe to the tenor and substance of this section suggesting such importance for genes and gene mutations in the evolution of the dietary influence on health. I already mentioned the epigenetic evidence for long-term evolutionary change that does not depend on genetic mutation. In addition, there also are extensive data now existing to show that it is not the presence or absence of genes, good or bad, that actually determines health outcomes. We now know that it is the nutritional control of gene expression that matters far more. In our own work on experimental animals, we were able to demonstrate a remarkable ability of nutrition to control gene(s) responsible for development of experimental cancers—this is briefly summarized in our book, “The China Study”. I should also note that it was the protein of cow’s milk (casein) that we used to show this remarkable nutritional effect on gene control, thus leading to our seriously questioning cow’s milk and its products on many grounds as a serious health food—a point that Cordain and I share.
7. Page 7, column 2, paragraph 1. Cordain cites an estimate of 4-8.5% of total energy as animal food for chimpanzees, our nearest living non-human primate, more or less agreeing with his suggestion of 3-5% (mostly insects, etc.) in a presentation of his at Boulder, Colorado, a couple years ago. In either case, he notes that this proportion was established sometime before 2.6 million years ago prior to the introduction of stone tool technology, inferring that this estimate may not be relevant for present day human consideration because of the greatly increased intake of animal foods up to at least 50% of total energy intake during Paleolithic times. If the ‘blueprints’ for nutrient intakes were established before 2.6 MYA, then why are these estimates of 3-8.5% (3-5%) irrelevant?
8. Page 11, column 2, paragraph 2. Relying on the hypothesized effects of N-nitroso compounds on gut tissue as an explanation of large bowel cancer is too narrow a view. It ignores factors such as bile acid secretion, increased anaerobiosis, dietary fat, dietary protein and an array of antioxidants as important modifiers. The claim that vegetarians do not have lower cancer rates is an overstatement; many studies show that cancer rates are somewhat lower. But more to the point, about 90% of vegetarians still consume dairy products, among other animal based foods, thus minimizing the difference in nutritional composition from non-vegetarian diets. It is somewhat surprising to me that we even observed statistically significant deceases in disease rates with such small differences in nutrient composition.
Summary
Although there are many more statements in Cordain’s paper that I do not support, I hope that these comments are sufficient. As stated in the beginning, my philosophy of nutrition is substantially different from Professor Cordain if, in fact, he even has a philosophy of nutrition, given his very negative views of this field. I admit that my views represent a paradigm shift from the traditional views of nutrition, including many of my colleagues in the field, and it is this very different world view that prompts so much of my skepticism of Cordain’s observations and views.
My paradigm of nutrition assumes that a holistic food effect is far greater than a reductionist nutrient effect. Not only does this view shape my interpretation of research findings, it also challenges the design of experimental studies, the assessment of food and health policy, the very nature of science itself, and the choices of what to eat.
Briefly, I strongly disagree with Cordain’s proposal that 15% dietary protein is likely to be too low. I say that is likely to be too high, especially if animal based foods are required to reach this level. My recommendation is about 8-12% total protein, with all or virtually all of it being plant-based. With whole plant-based foods, it is not easy to go above this range without adding animal based foods or over consuming high protein plant-based foods like legumes.
I arrive at this estimate of 8-12% total protein first by considering a wide variety of research evidence. I then use this evidence to decide what the total diet should be for good health, not only based on the hypothetically isolated effects of protein but also because of the combined and highly integrated effects of all the chemical compounds of this food. This is a diet comprised of a variety of nutrient-rich plant-based whole foods, with very little or no added fat, sugar and salt. I then ask what is the likely protein content of this diet and this is generally in the range of 8-12%. I then find it quite remarkable that so many of the adverse effects of diets higher in protein begin to show their effects at about 10-12% total protein, as summarized in our book, “The China Study”. I also find it quite remarkable that for many years multiple panels who have considered protein requirements arrive at a level of about 6-8% being the protein ‘requirement’ to match nitrogen loss, and about 8-10% being the allowance to hypothetically insure protein adequacy for 98% of the population.
These are my main broad based concerns with Cordain’s observations and conclusions.
1. I don’t find his evolutionary theory arguments to be persuasive. First, researchers in his own field cite considerable evidence to the contrary to refute his arguments concerning the contemporary hunter-gatherer societies. Second, the evidence presented by Cordain and others does not establish the proportion of energy provided by animal based foods. Third, I believe that he has put far too much weight on gene mutation events to describe the evolutionary basis for a normal, healthy diet and has overlooked the highly significant roles of epigenetic and adaptive mechanisms to also account for these same effects. This suggests that diet-disease associations observed in contemporary times are far more meaningful than what might have occurred during evolutionary times—at least since the last 2.5 million years or so.
2. His reliance on reductionist research findings and theory is at odds with the nutritional effect as it exists in reality. This is a serious flaw.
3. He omits the overwhelming findings on the adverse health effects of dietary protein as it increases within the range of 10-20% total protein. These increases are, in reality, almost always provided by increased consumption of animal-based foods.
4. Although Professor Cordain likely will not know this, he and others should nonetheless know of the remarkable healing effects now being routinely accomplished by my clinician colleagues as well as told to me by those who change to a low protein, low fat, whole foods plant-based diet.
References
1. Bogert LJ, Briggs GM, Calloway DH. Nutrition and Physical Fitness, Ninth Edition. W.B. Sauders Company, Philadelphia, 1973.
2. Dyerberg J, Bang HO. A hypothesis on the development of acute myocardial infarction in Greenlanders. Scand J Clin Lab Invest Suppl. 1982;161:7-13.
3. Jenkins DJ, Wolever TM, Taylor RH, Barker H, Fielden H, Baldwin JM, Bowling AC, Newman HC, Jenkins AL, Goff DV. Glycemic index of foods: a physiological basis for carbohydrate exchange. Am J Clin Nutr. 1981 Mar;34(3):362-6.
4. No authors listed. Position paper on trans fatty acids. ASCN/AIN Task Force on Trans Fatty Acids. American Society for Clinical Nutrition and American Institute of Nutrition. Am J Clin Nutr. 1996 May;63(5):663-70.
5. National Academy of Sciences, Institute of Medicine. Letter Report on Dietary Reference Intakes for Trans Fatty Acids, 2002. http://www.iom.edu/CMS/5410.aspx.
6. http://www.mypyramid.gov/
7. Willett WC, Stampfer MJ. Rebuilding the food pyramid. Sci Am. 2003 Jan;288(1):64-71
8. Krauss RM, Eckel RH, Howard B, et al. Dietary Guidelines: revision 2000: A statement for healthcare professionals from the Nutrition Committee of the American Heart Association. Circulation. 2000 Oct 31;102(18):2284-99.
9. Blanck HM, Gillespie C, Serdula MK, Khan LK, Galusk DA, Ainsworth BE .Use of low-carbohydrate, high-protein diets among americans: correlates, duration, and weight loss. MedGenMed. 2006 Apr 5;8(2):5.
10. Ornish D. Dr. Dean Ornish’s Program for Reversing Heart Disease: The Only System Scientifically Proven to Reverse Heart Disease Without Drugs or Surgery. New York : Random House, 1990.
11. Nesse RM, Stearns SC, Omenn GS. Medicine needs evolution. Science 2006;311:1071.
12. Dobzhansky T. Am Biol Teacher. 1973 March; 35:125-129.
13. Cordain L, Eaton SB, Sebastian A, Mann N, Lindeberg S, Watkins BA, O’Keefe JH, Brand-Miller .Origins and evolution of the Western diet: health implications for the 21st century. Am J Clin Nutr. 2005 Feb;81(2):341-54.
14. Cordain L, Miller JB, Eaton SB, Mann N, Holt SH, Speth JD. Plant-animal subsistence ratios and macronutrient energy estimations in worldwide hunter-gatherer diets. Am J Clin Nutr. 2000 Mar;71(3):682-92.
15. Cordain L, Eaton SB, Brand Miller J, Mann N, Hill K. The paradoxical nature of hunter-gatherer diets: Meat based, yet non-atherogenic. Eur J Clin Nutr 2002; 56 (suppl 1):S42-S52.
16. Cordain L, Watkins BA, Mann NJ. Fatty acid composition and energy density of foods available to African hominids: evolutionary implications for human brain development. World Rev Nutr Diet 2001, 90:144-161.
17. Bravata DM, Sanders L, Huang J, Krumholz HM, Olkin I, Gardner CD, Bravata DM. Efficacy and safety of low-carbohydrate diets: a systematic review.
JAMA. 2003 Apr 9;289(14):1837-50.
18. St Jeor ST, Howard BV, Prewitt TE, Bovee V, Bazzarre T, Eckel RH et al.. Dietary protein and weight reduction: a statement for healthcare professionals from the Nutrition Committee of the Council on Nutrition, Physical Activity, and Metabolism of the American Heart Association. Circulation. 2001 Oct 9;104(15):1869-74
19. Wright JD, J Kennedy-Stephenson J, Wang CY, McDowell MA, Johnson CL, National Center for Health Statistics, CDC. Trends in intake of energy and macronutrients—United States, 1971-2000. JAMA. 2004;291:1193-1194.
20. McDowell M, Briefel R, Alaimo K, et al. Energy and macronutrient intakes of persons ages 2 months and over in the United States: Third National Health and Nutrition Examination Survey, Phase 1, 1988–91.Washington, DC: US Government Printing Office, Vital and Health Statistics; 1994. CDC publication No. 255.
21. Rudman D, DiFulco TJ, Galambos JT, Smith RB 3rd, Salam AA, Warren WD. Maximal rates of excretion and synthesis of urea in normal and cirrhotic subjects.
J Clin Invest. 1973 Sep;52(9):2241-9.
22. Speth JD, Spielmann KA. Energy source, protein metabolism, and hunter-gatherer subsistence strategies. J Anthropological Archaeology 1983;2:1-31.
23. Speth JD. Early hominid hunting and scavenging: the role of meat as an energy source. J Hum Evol 1989;18:329-43.
24. Noli D, Avery G. Protein poisoning and coastal subsistence. J Archaeological Sci 1988;15:395-401.
25. Lieb CW. The effects on human beings of a twelve months’ exclusive meat diet. JAMA 1929;93:20-22.
26. Ogden CL, Fryar CD, Carroll MD, Flegal KM. Mean body weight, height and body mass index, United States 1960—2002 . Center for Disease Control. Advance Data from Vital and Health Statistics, No. 347, October 27, 2004.
27. Oliver WJ, Cohen EL, Neel JV. Blood pressure, sodium intake, and sodium related hormones in the Yanomamo Indians, a “no-salt” culture. Circulation. 1975 Jul;52(1):146-51.
28. Semaw S, Rogers MJ, Quade J, Renne PR, Butler RF, Dominguez-Rodrigo M, Stout D, Hart WS, Pickering T, Simpson SW. 2.6-Million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar, Ethiopia. J Hum Evol. 2003 Aug;45(2):169-77.
29. Bunn, HT, Kroll EM. Systematic butchery by Plio-Pleistocene hominids at Olduvai Gorge, Tanzania. Curr Anthropol 1986;20:365–398.
30. de Heinzelin J, Clark JD, White T, Hart W, Renne P, WoldeGabriel G, Beyene Y, Vrba E. Environment and behavior of 2.5-million-year-old Bouri hominids.
Science. 1999 Apr 23;284(5414):625-9
31. Asfaw B. White T, Lovejoy O, Latimer B, Simpson S, Suwa, G. Australopithecus garhi: A new species of early hominid from Ethiopia. Science 1999; 284, 629–635.
32. Vekua A, Lordkipanidze D, Rightmire GP, Agusti J, Ferring R, Maisuradze G, Mouskhelishvili A, Nioradze M, De Leon MP, Tappen M, Tvalchrelidze M, Zollikofer C. A new skull of early Homo from Dmanisi, Georgia. Science. 2002 Jul 5;297(5578):85-9.
33. Zhu RX, Potts R, Xie F. Hoffman KA, Deng CL, Shi CD, Pan YX, Wang HQ, Shi, RP, Wang YC, Shi GH, Wu NQ. New evidence on the earliest human presence at high northern latitudes in northeast Asia. Nature 2004; 431: 559–562.
34. Aiello LC, Wheeler P. The expensive tissue hypothesis. Curr Anthropol 1995; 36:199–222.
35. Leonard W.R, Robertson ML. Evolutionary perspectives on human nutrition: The influence of brain and body size on diet and metabolism. Am J Hum Biol 1994; 6: 77–88.
36. Pawlosky R., Barnes A., Salem, N. Essential fatty acid metabolism in the feline: Relationship between liver and brain production of long-chain polyunsaturated fatty acids. J Lipid Res 1994;35: 2032–2040.
37. Hussein N, Ah-Sing E, Wilkinson P, Leach C, Griffin BA, Millward DJ. Long-chain conversion of [13C] linoleic acid and alpha-linolenic acid in response to marked changes in their dietary intake in men. J Lipid Res. 2005 Feb;46(2):269-80.
38. Sturman JA, Hepner GW, Hofmann AF, Thomas PJ. Metabolism of [35S] taurine in man. J Nutr. 1975 Sep;105(9):1206-14.
39. Chesney RW, Helms RA, Christensen M, Budreau AM, Han X, Sturman JA. The role of taurine in infant nutrition. Adv Exp Med Biol. 1998;442:463-76.
40. Laidlaw SA, Shultz TD, Cecchino JT, Kopple JD. Plasma and urine taurine levels in vegans. Am J Clin Nutr. 1988 Apr;47(4):660-3.
41. Knopf K, Sturman JA, Armstrong M, Hayes KC. 1978. Taurine: An essential nutrient for the cat. J Nutr 1978;108: 773–778.
42. MacDonald ML, Rogers QR, Morris JG. Nutrition of the domestic cat, a mammalian carnivore. Annu Rev Nutr 1984; 4: 521–562.
43. Fam AG. Gout: excess calories, purines, and alcohol intake and beyond. Response to a urate-lowering diet. J Rheumatol. 2005 May;32(5):773-7.
44. Matzkies F, Berg G, Madl H. The uricosuric action of protein in man. Adv Exp Med Biol 1980;122A:227-31.
45. Loffler W. Grobner W, Medina R, Zollner N. Influence of dietary purines on pool size, turnover, and excretion of uric acid during balance conditions. Isotope studies using 15N-uric acid. Res Exp Med (Berl). 1982(2):113-123.
46. Oda M, Satta Y, Takenaka O, Takahata N. Loss of urate oxidase activity in hominoids and its evolutionary implications. Mol Biol Evol. 2002 May; 19(5): 640-53.
47. Abadeh S, Killacky J, Benboubetra M, Harrison R. Purification and partial characterization of xanthine oxidase from human milk. Biochim Biophys Acta. 1992 Jul 21;1117(1):25-32
48. Xu P, LaVallee P, Hoidal JR. Repressed expression of the human xanthine oxidoreductase gene. E-box and TATA-like elements restrict ground state transcriptional activity. J Biol Chem. 2000 Feb 25;275(8):5918-26.
49. Dessein PH, Shipton EA, Stanwix AE, Joffe BI, Ramokgadi J. Beneficial effects of weight loss associated with moderate calorie/carbohydrate restriction, and increased proportional intake of protein and unsaturated fat on serum urate and lipoprotein levels in gout: a pilot study. Ann Rheum Dis. 2000 Jul;59(7):539-43.
50. Teleki G. The omnivorous chimpanzee. Sci Am 1973; 228: 33–42.
51. Stanford CB. The hunting ecology of wild chimpanzees: Implications for the evolutionary ecology of Pliocene hominids. Am Anthropol 1996; 98: 96–113.
52. Schoeninger MJ, Moore J, Sept JM. Subsistence strategies of two “savanna” chimpanzee populations: The stable isotope evidence. Am J Primatol 1999: 49: 297–314.
53. van der Merwe NJ, Thackeray JF, Lee-Thorp JA, Luyt J. The carbon isotope ecology and diet of Australopithecus africanus at Sterkfontein, South Africa J Hum Evol 2003;44: 581–597.
54. Lee-Thorp J, Thackeray JF, van der Merwe N. The hunters and the hunted revisited. J Hum Evol 2000; 39: 565–576.
55. Sponheimer M, Lee-Thorp JA.. Differential resource utilization by extant great apes and australopithecines: Towards solving the C4 conundrum. Comp Biochem Physiol A 2003;136: 27–34.
56. Sussman RW. Foraging patterns of nonhuman primates and the nature of food preferences in man. Fed Proc 1978;37: 55–60.
57. Richards MP, Pettitt PB, Trinkaus E, Smith FH, Paunovic M, Karavanic, I. Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes. Proc Natl Acad Sci 2000;97: 7663–7666.
58. Bocherens H, Drucker DG, Billiou D, Patou-Mathis M, Vandermeersch B. Isotopic evidence for diet and subsistence pattern of the Saint-Cesaire I Neanderthal: review and use of a multi-source mixing model. J Hum Evol. 2005 Jul;49(1):71-87
59. Balter V, Simon L. Diet and behavior of the Saint-Cesaire Neanderthal inferred from biogeochemical data inversion. J Hum Evol. 2006 Oct;51(4):329-38.
60. Currat M, Excoffier L. Modern humans did not admix with Neanderthals during their range expansion into Europe. PLoS Biol. 2004 Dec;2(12):e421. Epub 2004 Nov 30
61. Richards MP, Hedges REM, Jacobi R, Current, A, Stringer C. Focus: Gough’s Cave and Sun Hole Cave human stable isotope values indicate a high animal protein diet in the British Upper Palaeolithic. J Archaeol Sci 2000;27: 1–3.
62. Gray JP. A corrected ethnographic atlas. World Cult J 1999;10: 24–85.
63. Hayden B. Subsistence and ecological adaptations of modern hunter/gatherers. In: RSO Harding, RSO, Teleki G. (Eds.), Omnivorous Primates. Columbia University Press, New York, 1981, pp. 344–421.
64. Kaplan H, Hill K, Lancaster J, Hurtado AM. A theory of human life history evolution: diet, intelligence, and longevity. Evol. Anthropol 2000;9:156–185.
65. Sempos CT, Liu K, Ernst ND. Food and nutrient exposures: what to consider when evaluating epidemiologic evidence. Am J Clin Nutr. 1999 Jun;69(6):1330S-1338S.
66. Potischman N, Weed DL. Causal criteria in nutritional epidemiology.
Am J Clin Nutr. 1999 Jun;69(6):1309S-1314S.
67. Freudenheim JL. Study design and hypothesis testing: issues in the evaluation of evidence from research in nutritional epidemiology. Am J Clin Nutr. 1999 Jun; 69(6): 1315S-1321S.
68. Fraser GE. A search for truth in dietary epidemiology. Am J Clin Nutr. 2003 Sep;78(3 Suppl):521S-525S.
69. Flegal KM. Evaluating epidemiologic evidence of the effects of food and nutrient exposures. Am J Clin Nutr. 1999 Jun;69(6):1339S-1344S.
70. Hu FB, Stampfer MJ, Manson JE, Rimm E, Colditz GA, Speizer FE, Hennekens CH, Willett WC. Dietary protein and risk of ischemic heart disease in women.
Am J Clin Nutr. 1999 Aug;70(2):221-7.
71. Campbell TC, Junshi C. Diet and chronic degenerative diseases: perspectives from China. Am J Clin Nutr. 1994 May;59(5 Suppl):1153S-1161S.
72. Campbell TC, Parpia B, Chen J. Diet, lifestyle, and the etiology of coronary artery disease: the Cornell China study. Am J Cardiol. 1998 Nov 26;82(10B):18T-21T.
73. Popkin BM. Where’s the fat? Trends in U.S. Diets 1965-1996. Prev Med 2001;32:245-54.
74. O’Dea K, Traianedes K, Chisholm K, Leyden H, Sinclair AJ. Cholesterol-lowering effect of a low-fat diet containing lean beef is reversed by the addition of beef fat. Am J Clin Nutr. 1990 Sep;52(3):491-4.
75. Wolfe BM & Giovannetti PM (1991): Short term effects of substituting protein for carbohydrate in the diets of moderately hypercholesterolemic human subjects. Metabolism 40, 338-343.
76. Wolfe BM & Piche LA (1999): Replacement of carbohydrate by protein in a conventional-fat diet reduces cholesterol and triglyceride concentrations in healthy normolipidemic subjects. Clin. Invest. Med. 22, 140-148.
77. O’Dea K (1984): Marked improvement in carbohydrate and lipid metabolism in diabetic Australian Aborigines after temporary reversion to traditional lifestyle. Diabetes 33, 596-603.
78. O’Dea K, Traianedes K, Ireland P, Niall M, Sadler J, Hopper J & DeLuise M (1989): The effects of diet differing in fat, carbohydrate, and fiber on carbohydrate and lipid metabolism in type II diabetes. J. Am. Diet. Assoc. 89, 1076-1086.
79. Layman DK, Boileau RA, Erickson DJ, Painter JE, Shiue H, Sather C, Christou DD. A reduced ratio of dietary carbohydrate to protein improves body composition and blood lipid profiles during weight loss in adult women. J Nutr. 2003 Feb;133(2):411-7.
80. Noakes M, Keogh JB, Foster PR, Clifton PM. Effect of an energy-restricted, high-protein, low-fat diet relative to a conventional high-carbohydrate, low-fat diet on weight loss, body composition, nutritional status, and markers of cardiovascular health in obese women. Am J Clin Nutr. 2005 Jun;81(6):1298-306.
81. Farnsworth E, Luscombe ND, Noakes M, Wittert G, Argyiou E, Clifton PM. Effect of a high-protein, energy-restricted diet on body composition, glycemic control, and lipid concentrations in overweight and obese hyperinsulinemic men and women.
Am J Clin Nutr. 2003 Jul;78(1):31-9.
82. Luscombe-Marsh ND, Noakes M, Wittert GA, Keogh JB, Foster P, Clifton PM.
Carbohydrate-restricted diets high in either monounsaturated fat or protein are equally effective at promoting fat loss and improving blood lipids. Am J Clin Nutr. 2005 Apr;81(4):762-72
83. Aude YW, Agatston AS, Lopez-Jimenez F, Lieberman EH, Marie Almon, Hansen M, Rojas G, Lamas GA, Hennekens CH. The national cholesterol education program diet vs a diet lower in carbohydrates and higher in protein and monounsaturated fat: a randomized trial. Arch Intern Med. 2004 Oct 25;164(19):2141-6.
84. McAuley KA, Hopkins CM, Smith KJ, McLay RT, Williams SM, Taylor RW, Mann JI. Comparison of high-fat and high-protein diets with a high-carbohydrate diet in insulin-resistant obese women. Diabetologia. 2005 Jan;48(1):8-16.
85. Appel LJ, Sacks FM, Carey VJ, Obarzanek E, Swain JF, Miller ER 3rd, Conlin PR, Erlinger TP, Rosner BA, Laranjo NM, Charleston J, McCarron P, Bishop LM; OmniHeart Collaborative Research Group. Effects of protein, monounsaturated fat, and carbohydrate intake on blood pressure and serum lipids: results of the OmniHeart randomized trial. JAMA. 2005 Nov 16;294(19):2455-64.
86. Kalopissis, AD Griffaton G, Fau D. Inhibition of hepatic very low density lipoprotein secretion on obese Zucker rats adapted to a high protein diet. Metabolism 1995;44:19-29.
87. Appel LJ. The effects of protein intake on blood pressure and cardiovascular disease. Curr Opin Lipidol. 2003 Feb;14(1):55-9. Review.
88. Elliott P. Protein intake and blood pressure in cardiovascular disease.
Proc Nutr Soc. 2003 May;62(2):495-504.
89. He J, Whelton PK. Elevated systolic blood pressure as a risk factor for cardiovascular and renal disease. J Hypertens Suppl. 1999 Jun;17(2):S7-13.
90. Burke V, Hodgson JM, Beilin LJ, Giangiulioi N, Rogers P, Puddey IB.
Dietary protein and soluble fiber reduce ambulatory blood pressure in treated hypertensives. Hypertension. 2001 Oct;38(4):821-6.
91. Washburn S, Burke GL, Morgan T, Anthony M. Effect of soy protein supplementation on serum lipoproteins, blood pressure, and menopausal symptoms in perimenopausal women. Menopause. 1999 Spring;6(1):7-13.
92. He J, Gu D, Wu X, Chen J, Duan X, Chen J, Whelton PK. Effect of soybean protein on blood pressure: a randomized, controlled trial.Ann Intern Med. 2005 Jul 5;143(1):1-9.
93. Hodgson JM, Burke V, Beilin LJ, Puddey IB. Partial substitution of carbohydrate intake with protein intake from lean red meat lowers blood pressure in hypertensive persons. Am J Clin Nutr. 2006 Apr;83(4):780-7
94. Nuttall FQ, Gannon MC. The metabolic response to a high-protein, low-carbohydrate diet in men with type 2 diabetes mellitus. Metabolism. 2006 Feb;55(2):243-51.
95. Nuttall FQ, Gannon MC. Metabolic response of people with type 2 diabetes to a high protein diet. Nutr Metab (Lond). 2004 Sep 13;1(1):6
52.
96. McAuley KA, Smith KJ, Taylor RW, McLay RT, Williams SM, Mann JI. Long-term effects of popular dietary approaches on weight loss and features of insulin resistance. Int J Obes (Lond). 2006 Feb;30(2):342-9.
97. Weigle DS, Breen PA, Matthys CC, Callahan HS, Meeuws KE, Burden VR, Purnell JQ. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am J Clin Nutr. 2005 Jul;82(1):41-8
98. Due A, Toubro S, Skov AR, Astrup A. Effect of normal-fat diets, either medium or high in protein, on body weight in overweight subjects: a randomised 1-year trial.
Int J Obes Relat Metab Disord. 2004 Oct;28(10):1283-90
99. Westerterp-Plantenga MS, Lejeune MP, Nijs I, van Ooijen M, Kovacs EM. High protein intake sustains weight maintenance after body weight loss in humans. Int J Obes Relat Metab Disord. 2004 Jan;28(1):57-64.
100. Lejeune MP, Kovacs EM, Westerterp-Plantenga MS. Additional protein intake limits weight regain after weight loss in humans. Br J Nutr. 2005 Feb;93(2):281-9.
101. Porrini M, Santangelo A, Crovetti R, Riso P, Testolin G, Blundell JE.
Reid M, Hetherington M. Relative effects of carbohydrates and protein on satiety -- a review of methodology. Neurosci Biobehav Rev 1997 May;21(3):295-308
102. Poppitt SD, McCormack D, Buffenstein R. Short-term effects of macronutrient preloads on appetite and energy intake in lean women. Physiol Behav 1998 Jun 1;64(3):279-85
103. Batterham RL, Heffron H, Kapoor S, Chivers JE, Chandarana K, Herzog H, Le Roux CW, Thomas EL, Bell JD, Withers DJ. Critical role for peptide YY in protein-mediated satiation and body-weight regulation. Cell Metab. 2006 Sep;4(3):223-33.
104. O’Brien KO, Abrams SA, Liang LK, Ellis KJ, Gagel RF. Increased efficiency of calcium absorption during short periods of inadequate calcium intake in girls. Am J Clin Nutr. 1996 Apr;63(4):579-83
105. Dawson-Hughes B, Harris SS, Rasmussen H, Song L, Dallal GE. Effect of dietary protein supplements on calcium excretion in healthy older men and women. J Clin Endocrinol Metab. 2004 Mar;89(3):1169-73
106. Spencer H, Kramer L, Osis D, Norris C. Effect of a high protein (meat) intake on calcium metabolism in man. Am J Clin Nutr. 1978 Dec;31(12):2167-80
107. Roughead ZK, Johnson LK, Lykken GI, Hunt JR. Controlled high meat diets do not affect calcium retention or indices of bone status in healthy postmenopausal women.
J Nutr. 2003 Apr;133(4):1020-6
108. Arjmandi BH, Khalil DA, Smith BJ, Lucas EA, Juma S, Payton ME, Wild RA.
Soy protein has a greater effect on bone in postmenopausal women not on hormone replacement therapy, as evidenced by reducing bone resorption and urinary calcium excretion. J Clin Endocrinol Metab. 2003 Mar;88(3):1048-54
109. Pannemans DL, Schaafsma G, Westerterp KR. Calcium excretion, apparent calcium absorption and calcium balance in young and elderly subjects: influence of protein intake. Br J Nutr. 1997 May;77(5):721-9.
110. Kerstetter JE, O’Brien KO, Insogna KL. Dietary protein affects intestinal calcium absorption. Am J Clin Nutr. 1998 Oct;68(4):859-65.
111. Kerstetter JE, O’Brien KO, Caseria DM, Wall DE, Insogna KL. The impact of dietary protein on calcium absorption and kinetic measures of bone turnover in women.
J Clin Endocrinol Metab. 2005 Jan;90(1):26-31.
112. Kerstetter JE, Wall DE, O’Brien KO, Caseria DM, Insogna KL. Meat and soy protein affect calcium homeostasis in healthy women. J Nutr. 2006 Jul;136(7):1890-5
113. Brenner BM, Meyer TW, Hostetter TH. Dietary protein intake and the progressive nature of kidney disease: the role of hemodynamically mediated glomerular injury in the pathogenesis of progressive glomerular sclerosis in aging, renal ablation, and intrinsic renal disease.N Engl J Med. 1982 Sep 9;307(11):652-9.
114. Lippert J, Ritz E, Schwarzbeck A, Schneider P. The rising tide of endstage renal failure from diabetic nephropathy type II--an epidemiological analysis.
Nephrol Dial Transplant. 1995;10(4):462-7.
115. Ritz E, Rychlik I, Locatelli F, Halimi S. End-stage renal failure in type 2 diabetes: A medical catastrophe of worldwide dimensions. Am J Kidney Dis. 1999 Nov;34(5):795-808.
116. Wrone EM, Carnethon MR, Palaniappan L, Fortmann SP; Third National Health and Nutrition Examination Survey. Association of dietary protein intake and microalbuminuria in healthy adults: Third National Health and Nutrition Examination Survey. Am J Kidney Dis. 2003 Mar;41(3):580-7.
117. Johnson DW. Dietary protein restriction as a treatment for slowing chronic kidney disease progression: the case against. Nephrology. 2006 Feb;11(1):58-62.
118. Skov AR, Toubro S, Bulow J, Krabbe K, Parving HH, Astrup A. Changes in renal function during weight loss induced by high vs low-protein low-fat diets in overweight subjects. Int J Obes Relat Metab Disord. 1999 Nov;23(11):1170-7.
119. Bingham SA. High-meat diets and cancer risk. Proc Nutr Soc. 1999 May;58(2):243-8.
120. Truswell AS. Meat consumption and cancer of the large bowel. Eur J Clin Nutr. 2002 Mar;56 Suppl 1:S19-24.
121. Larsson SC, Wolk A. Meat consumption and risk of colorectal cancer: A meta-analysis of prospective studies.Int J Cancer. 2006 Sep 21; [Epub ahead of print]
122. Bingham SA, Hughes R, Cross AJ. Effect of white versus red meat on endogenous N-nitrosation in the human colon and further evidence of a dose response. J Nutr. 2002 Nov;132(11 Suppl):3522S-3525S
123. Lunn JC, Kuhnle G, Mai V, Frankenfeld C, Shuker DE, Glen RC, Goodman JM, Pollock JR, Bingham SA. The effect of haem in red and processed meat on the endogenous formation of N-nitroso compounds in the upper gastrointestinal tract. Carcinogenesis. 2006 Oct 19; [Epub ahead of print].
124. Haorah J, Zhou L, Wang X, Xu G, Mirvish SS. Determination of total N-nitroso compounds and their precursors in frankfurters, fresh meat, dried salted fish, sauces, tobacco, and tobacco smoke particulates. J Agric Food Chem. 2001 Dec;49(12):6068-78
125. Gerrior S, Bente I. 2002. Nutrient Content of the U.S. Food Supply, 1909-99: A Summary Report.
U.S.D.A, Center for Nutrition Policy and Promotion. Home Economics Research Report No. 55.
126. Hughes R, Pollock JR, Bingham S. Effect of vegetables, tea, and soy on endogenous N-nitrosation, fecal ammonia, and fecal water genotoxicity during a high red meat diet in humans. Nutr Cancer. 2002;42(1):70-7.
127. Stefansson V. Cancer: Disease of Civilization. Hill and Wang, New York, 1960.
128. Patnaik AK, Liu SK, Johnson GF. Feline intestinal adenocarcinoma. A clinicopathologic study of 22 cases. Vet Pathol. 1976;13(1):1-10
129. Anthony JC, Lang CH, Crozier SJ, Anthony TG, MacLean DA, Kimball SR, Jefferson LS. Contribution of insulin to the translational control of protein synthesis in skeletal muscle by leucine. Am J Physiol Endocrinol Metab. 2002;282:E1092-101.
130. Layman DK. Role of leucine in protein metabolism during exercise and recovery. Can J Appl Physiol 2002;27:646-62.
131. Rasmussen BB, Tipton KD, Miller SL, Wolf SE, Wolfe RR. An oral essential amino acid-carbohydrate supplement enhances muscle protein anabolism after resistance exercise.J Appl Physiol. 2000 Feb;88(2):386-92.
132. Tipton KD, Rasmussen BB, Miller SL, Wolf SE, Owens-Stovall SK, Petrini BE, Wolfe RR. Timing of amino acid-carbohydrate ingestion alters anabolic response of muscle to resistance exercise. Am J Physiol Endocrinol Metab. 2001 Aug;281(2):E197-206.
133. Levenhagen DK, Gresham JD, Carlson MG, Maron DJ, Borel MJ, Flakoll PJ. Postexercise nutrient intake timing in humans is critical to recovery of leg glucose and protein homeostasis. Am J Physiol Endocrinol Metab. 2001;280:E982-93.
134. Blomstrand E. Amino acids and central fatigue. Amino Acids 2001;20:25-34.
135. Kuhn T. The Structure of Scientific Revolutions. University of Chicago Press, Chicago, 1970.
136. Nishikimi M, Yagi K. Molecular basis for the deficiency in humans of gulonolactone oxidase, a key enzyme for ascorbic acid biosynthesis. Am J Clin Nutr. 1991 Dec;54(6 Suppl):1203S-1208S.
137. Rand WM, Pellett PL, Young VR. Meta-analysis of nitrogen balance studies for estimating protein requirements in healthy adults. Am J Clin Nutr. 2003 Jan;77(1):109-27.
138. Vieth R. Why the optimal requirement for Vitamin D3 is probably much higher than what is officially recommended for adults. J Steroid Biochem Mol Biol. 2004 May;89-90(1-5):575-9.
139. Dawson-Hughes B, Heaney RP, Holick MF, Lips P, Meunier PJ, Vieth R. Estimates of optimal vitamin D status. Osteoporos Int. 2005 Jul;16(7):713-6.
140. Miller M. Niacin as a component of combination therapy for dyslipidemia.
Mayo Clin Proc. 2003 Jun;78(6):735-42.
141. Chittenden RH. Physiologic economy in nutrition. New York: Heinemann, 1905.
142. Remer T, Manz F. Potential renal acid load of foods and its influence on urine pH. J Am Diet Assoc. 1995 Jul;95(7):791-7.
143. Cordain L Cereal grains: humanity’s double-edged sword. World Rev Nutr Diet. 1999;84:19-73.
144. Berlyne GM, Ben Ari J, Nord E, Shainkin R: Bedouin osteomalacia due to calcium deprivation caused by high phytic acid content of unleavened bread. Am J Clin Nutr 1973;26:910-11.
145. Ford JA, Colhoun EM, McIntosh WB, Dunnigan MG: Biochemical response of late rickets and osteomalacia to a chupatty-free diet. Brit Med J 1972;2:446-47.
146. Robertson I, Ford JA, McIntosh WB, Dunnigan MG: The role of cereals in the aetiology of nutritional rickets: the lesson of the Irish national nutritional survey 1943-8. Brit J Nutr 1981;45:17-22.
147. Stephens WP, Berry JL, Klimiuk PS, Mawer EB: Annual high dose vitamin D prophylaxis in asian immigrants. Lancet 1981;2:1199-201.
148. Ewer TK: Rachitogenicity of green oats. Nature 1950;166:732-33.
149. MacAuliffe T, Pietraszek A, McGinnis J: Variable rachitogenic effects of grain and alleviation by extraction or supplementation with vitamin D, fat and antibiotics. Poultry Sci 1976;55:2142-47.
150. Hidiroglou M, Ivan M, Proulx JG, Lessard JR: Effect of a single intramuscular dose of vitamin D on concentrations of liposoluble vitamins in the plasma of heifers winter-fed oat silage, grass silage or hay. Can J Anim Sci 1980;60:311-18.
151. Sly MR, van der Walt WH, Du Bruyn DB, Pettifor JM, Marie PJ: Exacerbation of rickets and osteomalacia by maize: a study of bone histomorphometry and composition in young baboons. Calcif Tissue Int 1984;36:370-79.
152. Batchelor AJ, Compston JE: Reduced plasma half-life of radio-labelled 25-hydroxyvitamin D3 in subjects receiving a high fiber diet. Brit J Nutr 1983;49:213-16.
153. Clements MR, Johnson L, Fraser DR: A new mechanism for induced vitamin D deficiency in calcium deprivation. Nature 1987;325:62-65.
154. Navarro A, Diaz MP, Munoz SE, Lantieri MJ, Eynard AR. Characterization of meat consumption and risk of colorectal cancer in Cordoba, Argentina.Nutrition. 2003 Jan;19(1):7-10.
155. Muscat JE, Wynder EL. The consumption of well-done red meat and the risk of colorectal cancer. Am J Public Health. 1994 May;84(5):856-8.
156. Sinha R, Chow WH, Kulldorff M, Denobile J, Butler J, Garcia-Closas M, Weil R, Hoover RN, Rothman N. Well-done, grilled red meat increases the risk of colorectal adenomas. Cancer Res. 1999 Sep 1;59(17):4320-4
157. Williams JH, Phillips TD, Jolly PE, Stiles JK, Jolly CM, Aggarwal D Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. Am J Clin Nutr. 2004 Nov;80(5):1106-22.
158. Dunaif GE, Campbell TC. Dietary protein level and aflatoxin B1-induced preneoplastic hepatic lesions in the rat. J Nutr. 1987 Jul;117(7):1298-302.
159. Dunaif GE, Campbell TC. Relative contribution of dietary protein level and aflatoxin B1 dose in generation of presumptive preneoplastic foci in rat liver.
J Natl Cancer Inst. 1987 Feb;78(2):365-9.
160. Horio F, Youngman LD, Bell RC, Campbell TC. Thermogenesis, low-protein diets, and decreased development of AFB1-induced preneoplastic foci in rat liver.
Nutr Cancer. 1991;16(1):31-41.
161. Youngman LD, Campbell TC. The sustained development of preneoplastic lesions depends on high protein intake. Nutr Cancer. 1992;18(2):131-42.
162. Fasano A, Berti I, Gerarduzzi T, Not T, Colletti RB, Drago S, Elitsur Y, Green PH, Guandalini S, Hill ID, Pietzak M, Ventura A, Thorpe M, Kryszak D, Fornaroli F, Wasserman SS, Murray JA, Horvath K. Prevalence of celiac disease in at-risk and not-at-risk groups in the United States: a large multicenter study.Arch Intern Med. 2003 Feb 10;163(3):286-92.
163. Cordain L, Toohey L, Smith MJ, Hickey MS. Modulation of immune function by dietary lectins in rheumatoid arthritis. Br J Nutr. 2000 Mar;83(3):207-17.
164. Key TJ, Fraser GE, Thorogood M, Appleby PN, Beral V, Reeves G, Burr ML, Chang-Claude J, Frentzel-Beyme R, Kuzma JW, Mann J, McPherson K. Mortality in vegetarians and nonvegetarians: detailed findings from a collaborative analysis of 5 prospective studies. Am J Clin Nutr. 1999 Sep;70(3 Suppl):516S-524S
Protein plays a litany of roles in living systems: structural elements, peptide hormones, cell recognition, antibodies… the list is staggering and continues to grow as our understanding of biology expands. What, however, is the role of dietary protein in health and disease in humans? Is the source, type and quantity intimately and directly tied to optimal physical development and continued wellbeing? Is it causative or preventative of disease? How do we know, and how can we know?
One would think this question should be straightforward and easily answered; as you will soon see the question is anything but simple! In the pages that follow, two scientists at the top of their respective fields--Dr. T. Colin Campbell, Professor of Nutritional Biochemistry at Cornell University, author of The China Study and Dr. Loren Cordain Professor, Department of Health & Exercise Science, Colorado State University, author of The Paleo Diet—make their competing cases for the role of dietary protein in health and disease.
The Evolutionary Basis for the Therapeutic Effects of High Protein Diets
Loren Cordain, Ph.D, Professor
Department of Health and Exercise Science
Colorado State University
Introduction
Although humanity has been interested in diet and health for thousands of years, the organized, scientific study of nutrition has a relatively recent past. For instance, the world’s first scientific journal devoted entirely to diet and nutrition, The Journal of Nutrition only began publication in 1928. Other well known nutrition journals have a more recent history still: The British Journal of Nutrition (1947), The American Journal of Clinical Nutrition (1954), and The European Journal of Clinical Nutrition (1988). The first vitamin was “discovered” in 1912 and the last vitamin (B12) was identified in 1948 (1). The scientific notion that omega 3 fatty acids have beneficial health effects dates back only to the late 1970’s (2), and the characterization of the glycemic index of foods only began in 1981 (3).
Nutritional science is not only a newly established discipline, but it is also a highly fractionated, contentious field with constantly changing viewpoints on both major and minor issues that impact public health. For example, in 1996 a task force of experts from the American Society for Clinical Nutrition (ASCN) and the American Institute of Nutrition (AIN) came out with an official position paper on trans fatty acids stating, “We cannot conclude that the intake of trans fatty acids is a risk factor for coronary heart disease” (4).
Fast forward 6 short years to 2002 and the National Academy of Sciences, Institute of Medicine’s report on trans fatty acids (5) stating, “Because there is a positive linear trend between trans fatty acid intake and total and LDL (“bad”) cholesterol concentration, and therefore increased risk of cardiovascular heart disease, the Food and Nutrition Board recommends that trans fatty acid consumption be as low as possible while consuming a nutritionally adequate diet”.
These kinds of complete turnabouts and divergence of opinion regarding diet and health are commonplace in the scientific, governmental and medical communities. The official U.S. governmental recommendations for healthy eating are outlined in the “My Pyramid” program (6) which recently replaced the “Food Pyramid”, both of which have been loudly condemned for nutritional shortcomings by scientists from the Harvard School of Public Health (7). Dietary advice by the American Heart Association (AHA) to reduce the risk of coronary heart disease (CHD) is to limit total fat intake to 30% of total energy, to limit saturated fat to <10% of energy and cholesterol to <300 mg/day while eating at least 2 servings of fish per week (8). Although similar recommendations are proffered in the USDA “My Pyramid”, weekly fish consumption is not recommended because the authors of these guidelines feel there is only “limited” information regarding the role of omega 3 fatty acids in preventing cardiovascular disease (6). Surprisingly, the personnel makeup of both scientific advisory boards is almost identical. At least 30 million Americans have followed Dr. Atkins advice to eat more fat and meat to lose weight (9). In utter contrast, Dean Ornish tells us fat and meat cause cancer, heart disease and obesity, and that we would all would be a lot healthier if we were strict vegetarians (10). Who’s right and who’s wrong? How in the world can anyone make any sense out of this apparent disarray of conflicting facts, opinions and ideas?
In mature and well-developed scientific disciplines there are universal paradigms that guide scientists to fruitful end points as they design their experiments and hypotheses. For instance, in cosmology (the study of the universe) the guiding paradigm is the “Big Bang” concept showing that the universe began with an enormous explosion and has been expanding ever since. In geology, the “Continental Drift” model established that all of the current continents at one time formed a continuous landmass that eventually drifted apart to form the present-day continents. These central concepts are not theories for each discipline, but rather are indisputable facts that serve as orientation points for all other inquiry within each discipline. Scientists do not know everything about the nature of the universe, but it is absolutely unquestionable that it has been and is expanding. This central knowledge then serves as a guiding template that allows scientists to make much more accurate and informed hypotheses about factors yet to be discovered.
The study of human nutrition remains an immature science because it lacks a universally acknowledged unifying paradigm (11). Without an overarching and guiding template, it is not surprising that there is such seeming chaos, disagreement and confusion in the discipline. The renowned Russian geneticist Theodosius Dobzhansky (1900-1975) said, “Nothing in biology makes sense except in the light of evolution” (12). Indeed, nothing in nutrition seems to make sense because most nutritionists have little or no formal training in evolutionary theory, much less human evolution. Nutritionists face the same problem as anyone who is not using an evolutionary model to evaluate biology: fragmented information and no coherent way to interpret the data.
All human nutritional requirements like those of all living organisms are ultimately genetically determined. Most nutritionists are aware of this basic concept; what they have little appreciation for is the process (natural selection) which uniquely shaped our species’ nutritional requirements. By carefully examining the ancient environment under which our genome arose, it is possible to gain insight into our present day nutritional requirements and the range of foods and diets to which we are genetically adapted via natural selection (13-16). This insight can then be employed as a template to organize and make sense out of experimental and epidemiological studies of human biology and nutrition (11).
The Dietary Protein Conundrum: How Much is Enough?
An important dietary issue that has come under debate in recent years is the safety of high protein diets and their long term influence upon health and well being (17, 18). In the current U.S. diet the average protein intake is 98.6 g/day (15.5 % of total energy) for men and 67.5 g/day (15.1 % of total energy) for women (19). Animal products provide approximately 75 % of the protein in the U.S. food supply followed by dairy, cereals, eggs, legumes, fruits and vegetables (20). Diets containing 20 % or more of their total energy as protein have been labeled “high protein diets” and those containing 30% or more energy as protein have been dubbed “very high protein diets” (18). Accordingly, a “high protein diet” for the average U.S. male daily energy intake (2,618 kcal (19)) would contain between 125 to 186 grams of protein per day and for the average female (1,877 kcal (19)) between 89 to 133 grams of protein per day.
At this point, it should be noted that there is a physiological limit to the amount of protein that can be ingested before it becomes toxic (14, 21). A byproduct of dietary protein metabolism is nitrogen, which in turn is converted into urea by the liver and then excreted by the kidneys into the urine. The upper limit of protein ingestion is determined by the liver’s ability to synthesize urea. When nitrogen intake from dietary protein exceeds the ability of the liver to synthesize urea, excessive nitrogen (as ammonia) spills into the bloodstream causing hyperammonemia and toxicity (14, 21). Additionally excess amino acids from the metabolism of high amounts of dietary protein may become toxic by entering the circulation causing hyperaminoacidemia (14, 21).
The avoidance of the physiological effects of protein excess has been an important factor in shaping the subsistence strategies of hunter-gatherers (22- 24). Multiple historical and ethnographic accounts have documented the deleterious health effects that have occurred when humans were forced to rely solely upon the fat depleted, lean meat of wild animals (22). Excess consumption of dietary protein from the lean meats of wild animals leads to a condition referred to by early American explorers as “rabbit starvation” which initially results in nausea, then diarrhea and eventual death (22). Clinical documentation of this syndrome is virtually non-existent, except for a single case study (25).
Using known maximal rates of urea synthesis (MRUS) in normal subjects [65 mg N/h - kg (body weight )0.75 ] (range 55-76), it is possible to calculate the maximal protein intake, beyond which will exceed MRUS and result in hyperammonemia and hyperaminoacidemia (21). The mean maximal protein intake for the average weight U.S. male (189.4 lbs (26)) is then 270 g/day (range 233-322 g/day), and for an average weight female (162.8 lbs (26)), 246 g/day (range 208-288 g/day). Consequently, “very high protein diets” for the average U.S. male could range from 187 to 270 g/day and for females, 134 to 246 g/day.
So let’s summarize a few key points. The average protein intake in the U.S. is about 15 % of the normal daily caloric intake. Diets labeled as “high protein” contain 20-29 % protein of the normal daily caloric intake, and diets with 30-40 % protein are branded “very high protein”. It should be pointed out that this categorization is completely arbitrary and based almost entirely upon comparisons to the U.S. norm. A salient question from an evolutionary perspective would be, “Is the average U.S. protein intake necessarily average or normal for our species?” For example, blood pressure in the U.S. and most other westernized countries is considered “normal” when systolic pressure is 120 mm Hg and diastolic pressure is 80 mm Hg. However, in many non-westernized people these values would be higher than normal. Consider the data in Figure 1 below showing blood pressure in the Yanomamo Indians of Brazil, a non-salt consuming society. Not only is blood pressure lower than normal western values, but it stays uniform throughout life and does not rise with age (27).
Figure 1. Blood pressure in a group of 506 Brazilian Indians (26).
In order to objectively answer the question whether or not high protein diets have detrimental or therapeutic health effects compared to the U.S. norm (15 % total energy), it may be useful to frame this question in an evolutionary perspective before examining the experimental and epidemiological evidence.
High Protein Diets: The Evolutionary Evidence
The Fossil Evidence
A number of lines of evidence suggest that meat eating and high protein diets have been a component of human nutrition since the very origins of our genus Homo. Beginning approximately 2.6 million years ago (MYA), the hominin species that eventually led to Homo began to include more animal food in their diet. A number of lines of evidence support this viewpoint. First, the very first stone tools (Oldowan lithic technology) appear in the fossil record 2.6 MYA (28), and there is clear cut evidence to show that these tools were used to butcher and disarticulate animal carcasses (29, 30). Stone tool cut marks on the bones of prey animals and evidence for marrow extraction appear concurrently in the fossil record with the development of Oldowan lithic technology by at least 2.5 MYA (Figure 2) (30). It is not entirely clear which specific early hominin species or group of species manufactured and used these earliest of stone tools; however, Australopithecus garhi is a likely candidate (30, 31).
The development of stone tools and the increased dietary reliance on animal foods allowed early African hominins to colonize northern latitudes outside of Africa where plant foods would have been seasonally restricted. Early Homo skeletal remains and Oldowan lithic technology appear at the Dmanisi site in the Republic of Georgia (40° N) by 1.75 MYA (32), and more recently Oldowan tools dating to 1.66 MYA have been discovered at the Majuangou site in North China (40° N) (33). Both of these tool-producing hominins would likely have consumed considerably more animal food than pre-lithic hominins living in more temperate African climates, and it is likely the majority of their daily energy was obtained from animal foods during winter and early spring when plant food sources would have been scarce or unavailable.
Figure 2: The earliest evidence for meat and marrow extraction dating to 2.5 million years ago (30).
The Genetic Evidence
In addition to the fossil evidence suggesting a trend for increased animal food consumption, hominins may have experienced a number of genetic adaptations to animal-based diets early in our genus’s evolution analogous to those of obligate carnivores such as felines. Carnivorous diets reduce evolutionary selective pressures that act to maintain certain anatomical and physiological characteristics needed to process and metabolize high amounts of plant foods. In this regard, hominins, like felines, have experienced a reduction in gut size and metabolic activity along with a concurrent expansion of brain size and metabolic activity as they included more energetically dense animal food into their diets (16, 34, 35). Further, similar to obligate carnivores (36), humans maintain an inefficient ability to chain elongate and desaturate 18 carbon fatty acids to their product 20 and 22 carbon fatty acids (37). Since 20 and 22 carbon fatty acids are essential cellular lipids, then evolutionary reductions in desaturase and elongase activity in hominins indicate that preformed dietary 20 and 22 carbon fatty acids (found only in animal foods) were increasingly incorporated in lieu of their endogenously synthesized counterparts derived from 18 carbon plant fatty acids. Finally, our species has a limited ability to synthesize the biologically important amino acid, taurine, from precursor amino acids (38, 39), and vegetarian diets in humans result in lowered plasma and urinary concentrations of taurine (40). Like felines (41, 42) the need to endogenously synthesize taurine may have been evolutionarily reduced in humans because exogenous dietary sources of preformed taurine (found only in animal food) had relaxed the selective pressure formerly requiring the need to synthesize this conditionally essential amino acid.
Another genetic adaptation to a high meat diet involves the metabolism of purines. Purines are the nitrogenous base pairs which form the structural cross rung molecules of both DNA and RNA. As DNA and RNA are broken down within cells, the purines then can be metabolized into uric acid by the liver and a few other tissues within the body. The liver receives purines from two sources: 1) the diet, and 2) the daily breakdown of the body’s own tissues. About 2/3 of the daily purine load comes from the body’s turnover of cells, while 1/3 comes from the diet (43). When the combined purine load (from both diet and turnover of the body’s own cells) exceeds the kidney’s ability to excrete it, blood concentrations of uric acid rise, thereby increasing the risk for gout, a painful disease caused by formation of uric acid crystals in the joints. Although high protein, meat based diets contain high amounts of purines and would be expected to promote gout symptoms, protein ingestion actually decreases blood uric acid levels by increasing uric acid excretion (44). This seemingly paradoxical effect occurs because the kidney increases its excretion of uric acid when faced with elevated dietary purines (45). But more importantly, over the course of evolution, humans have evolved a genetic mutation which tends to prevent uric acid synthesis in the liver. Humans avoid the overproduction of uric acid in the face of increasing dietary purine intake from meats by decreasing the activity of an enzyme called xanthine oxidoreductase (46), a key catalyst in the final synthesis of uric acid. Compared to other animals, xanthine oxidase activity is almost 100 times lower in humans (47). This evolutionary adaptation has occurred because the gene coding for xanthine oxidoreductase has been repressed (48). The final proof of the pudding has been borne out by dietary interventions showing that high protein, low glycemic load diets actually normalized serum uric acid concentrations in 7 of 12 gout patients and significantly decreased gout attacks (49).
The Isotopic Fossil Evidence
Since the evolutionary split between hominins and pongids (apes) approximately 7 million years ago, the available evidence shows that all species of hominins ate an omnivorous diet composed of minimally processed, wild-plant, and animal foods. In support of this view is the omnivorous nature of chimpanzees, the closest living pongid link to hominins. Although chimpanzees (Pan paniscus and Pan troglodytes), our genetically closest nonhuman relatives, primarily consume a frugivorous diet, they still eat a substantial amount of meat obtained throughout the year from hunting and scavenging (50-52). Observational studies of wild chimpanzees demonstrate that during the dry season meat intake is about 65 g per day for adults (51). Accordingly, it is likely that the very earliest hominins would have been capable of obtaining animal food through hunting and scavenging in a manner similar to chimpanzees.
Carbon isotope data also support the notion that early hominins were omnivorous. By about 3 million years ago MYA Australopithecus africanus obtained a significant portion of food from C4 sources (grasses, particularly seeds and rhizomes; sedges; invertebrates, including locusts and termites; grazing mammals; and perhaps even insectivores and carnivores)(53). Other fossils of early African hominins, including Australopithecus robustus and Homo ergaster, maintain carbon isotope signatures characteristic of omnivores (54, 55). The finding of C4 in Australopithecus robustus fossils refutes the earlier view that this hominin was vegetarian (54).
There is little evidence to the contrary that animal foods have always played a significant role in the diets of all hominin species. Increased reliance on animal foods not only allowed for enhanced encephalization (brain expansion relative to body weight) and its concomitant behavioral sophistication (16, 34, 35), but this dietary practice also permitted colonization of the world outside of Africa. An unresolved issue surrounding hominin diets is the relative amounts of plant and animal foods that were typically consumed.
Before the advent of Oldowan lithic technology about 2.6 MYA quantitative estimates of hominin energy intake from animal food sources are unclear, other than they were likely similar to, or greater than, estimated values (4%–8.5% total energy) for chimpanzees (51, 56)). Although all available data point to increasing animal food consumption following the arrival of stone tool technology, the precise contribution of either animal or plant food to energy is unclear. Obviously, then as now, no single (animal/plant) subsistence ratio would have been necessarily representative of all populations or species of hominins. However, there are a number of lines of evidence which suggest more than half (>50%) of the average daily energy intake for most Paleolithic hominin species and populations of species was obtained from animal foods.
Richards, Pettitt, and colleagues (57) have examined stable isotopes (δ13C and δ15N) in two Neanderthal specimens (~28,000—29,000 years ago) from Vindija Cave in northern Croatia and contrasted these isotopic signatures to those in fossils of herbivorous and carnivorous mammals from the same ecosystem. The analysis demonstrated that Neanderthals, similar to wolves and arctic foxes, behaved as top-level carnivores, obtaining all of their protein from animal sources (57). More recent studies corroborate this earlier work and points to Neanderthals “as top predators in an open environment, with little variation through time and space” (58), and “the percentage of plants in the Neanderthal diet must have been close to zero (59).
Because Neanderthals were not direct predecessors of modern humans (60), it may be more relevant to examine the isotopic data from fully modern humans living during the Pleistocene. An analysis was made of five Upper Paleolithic Homo sapiens specimens dated to ~11,700–12,380 years ago from Gough’s and Sun Hole Caves in Britain (61). The data indicated these hunter-gatherers were consuming animal protein year-round at a higher trophic level than the artic fox.
All of these studies (57-62) could be criticized as not being representative of typical hominin diets, as these two species lived in climates and ecosystems that fostered an abundance of large, huntable mammals, which were preyed upon preferentially. Additional clues to the typical plant-to-animal subsistence ratio in Paleolithic hominin diets can be found in the foraging practices of historically studied hunter-gatherers.
The Ethnographic Evidence
Our analysis (Figure 3) of the Ethnographic Atlas data (62) showed that the dominant foods in the majority of historically studied hunter-gatherer diets were derived from animal food sources (14). Most (73%) of the world’s hunters-gatherers obtained >50 percent of their subsistence from hunted and fished animal foods, whereas only 14 percent of worldwide hunter gatherers obtained >50 percent of their subsistence from gathered plant foods. For all 229 hunter-gatherer societies, the median subsistence dependence on animal foods was 56 percent to 65 percent. In contrast, the median subsistence dependence on gathered plant foods was 26 percent to 35 percent (14).
The major limitation of ethnographic data is that the preponderance of it is subjective in nature, and the assigned scores for the five basic subsistence economies in the Ethnographic Atlas are not precise, but rather are approximations (63). Fortunately, more exact, quantitative dietary studies were carried out on a small percentage of the world’s hunter gatherer societies (15, 64). Table 1 lists these studies and shows the plant-to-animal subsistence ratios by energy. The average score for animal food subsistence is 65 percent, while that for plant-food subsistence is 35 percent. These values are similar to our analysis of the entire (n = 229) sample of hunter-gatherer societies listed in the Ethnographic Atlas in which the mean score for animal food subsistence was 68 percent and that for plant food was 32 percent (14). When the two polar hunter-gatherer populations, who have no choice but to eat animal food because of the inaccessibility of plant foods, are excluded from Table 1 the mean score for animal subsistence is 59 percent and that for plant-food subsistence is 41 percent. These animal-to-plant subsistence values fall within the same respective class intervals (56%–65% for animal food; 26%–35% for plant food) as those we estimated from the ethnographic data when the confounding influence of latitude was eliminated (14). Consequently, there is remarkably close agreement between the quantitative data in Table 1 and the ethnographic data (14) that animal food comprised more than half of the energy in historically studied hunter-gatherer diets.
Based upon hunter gatherer plant to animal subsistence ratios and the known macronutrient contents of wild plant and animal foods, it is possible to estimate the macronutrient content of these diets (14). The typical hunter-gatherer protein intake would have fallen between 19 and 35 % of total energy (14), values which would be labeled either “high” or “very high” protein diets when compared to current U.S. values (15 %). Consequently, when framed in an evolutionary context, current western dietary protein intakes fall outside the range of diets that would have conditioned the human genome for nearly 2 million years. The evolutionary template would then suggest that when dietary protein intakes are restored to levels that our species is genetically accustomed, good health will prevail. Conversely, lower or higher values likely result in ill health. Let’s see what the experimental evidence shows.
Figure 3. Frequency distribution of subsistence dependence upon animal foods in world wide hunter gatherer societies (n = 229).
Table 1. Quantitatively determined proportions of plant and animal food in hunter-gatherer diets (15, 64).
Dietary Protein: Health and Well Being
Establishing Cause and Effect between Diet and Disease
One of the challenges faced by nutritional scientists when they ultimately make recommendations regarding what we should and should not eat is to establish cause and effect between a dietary element and the subsequent development or prevention of disease. Some foods and some dietary habits promote good health whereas others promote disease. Figure 4 demonstrates the four primary procedures by which causality is established between diet and disease (65, 66).
No single procedure alone can establish cause and effect (65, 66), nor can any single study prove causality (67). Observational epidemiological studies can only show relationships among variables and are notorious for showing conflicting results (68 ) and cannot provide decisive evidence by themselves either for or against specific hypotheses (69). For example increased animal protein has been associated with a decreased risk for coronary heart disease (CHD) in a large group of nurses (The Nurses Health Study) (70), whereas exactly the opposite association was found for markers of CHD and meat consumption in people from rural China (71, 72). An analogy here may be appropriate to show you why observational epidemiological studies can only show relationships and not establish causality. In New York City, there is a strong association between the size of a structure fire and the number of fire trucks at the fire, but can we conclude that more fire trucks cause bigger fires?
In order to establish cause and effect between diet and disease, it takes more than just observational epidemiological evidence (69). There must also be what is referred to as “biological plausibility” in which evidence gathered from tissue, animal and short term human metabolic studies support causality (66). When observational epidemiological evidence is augmented by biological plausibility studies and confirmed by randomized controlled trials, the case for causality becomes ever more convincing. In regard to optimal amounts of dietary protein, the bulk of the evidence from tissue and animal studies and from human dietary interventions provides a compelling case for the therapeutic effects of high protein diets.
Figure 4. Analytical procedures for establishing cause and effect between diet and disease
Dietary Protein and Cardiovascular Disease
One of the reasons why observational epidemiological studies yield contradictory results is because of the influence of confounding variables which cause confusion in the interpretation of the results because of the mixing of effects from two or more variables (68). For example, although some observational studies have shown a positive association between animal protein and cardiovascular disease (CVD), it is entirely possible that this association is spurious because the measurement of animal protein is confounded by another variable that is also linked to CVD. Meat is a major source of animal protein in the U.S. diet (20), but it is also a major source of saturated fat (73). Because meat comes as an inseparable package of (protein + saturated fat), animal protein ingestion will be highly correlated to saturated fat, thereby making it difficult to disengage the atherogenic effect of saturated fat from that of animal protein. Accordingly, experimental studies are more useful to determine the true effect animal protein may have upon cardiovascular risk factors because they can be designed to isolate the protein effects from the saturated fat effects.
Sinclair and colleagues (74) performed an experiment in which they fed 10 adults a low fat, lean beef-based diet for 5 weeks. Energy intake was kept constant over the 5 week study. Total blood cholesterol concentrations fell significantly within 1 week of commencing the diet, but rose as beef fat drippings were added in a stepwise manner in weeks 4 and 5. The authors concluded, “. . . it is the beef fat, not lean beef itself, that is associated with elevations in cholesterol concentrations”.
Numerous short term human dietary interventions have demonstrated the therapeutic effect of lean, animal based protein upon blood lipid parameters. Wolfe and colleagues have shown that the isocaloric substitution of protein (23% energy) for carbohydrate in moderately hypercholesterolemic subjects resulted in significant decreases in total, LDL and VLDL cholesterol, and triglycerides while HDL cholesterol increased (75). Similar blood lipid changes have been observed in normal healthy subjects (76) and in type II diabetic patients in conjunction with improvements in glucose and insulin metabolism (77, 78). A litany of more recent studies has confirmed that elevations in dietary protein have a beneficial effect upon blood lipid profiles (79-85). The mechanism or mechanisms of action of high protein diets upon blood lipid chemistry are not clear; however animal studies suggest that the beneficial effects are caused by their powerful inhibition of hepatic VLDL synthesis, perhaps by altering apoprotein synthesis and assembly in the liver (86).
The relationship between protein intake and blood pressure has been comprehensively examined in observational population studies, which support the notion that higher protein intake can lower blood pressure (87-89). A substantial number of randomized controlled trials have demonstrated that higher dietary protein either from soy (90-92), mixed dietary sources (85) or from lean red meat (93) significantly lower blood pressure.
Dietary Protein and Insulin/Glucose Metabolism and Weight Regulation
In addition to reducing CVD risk by improving the blood lipid profile and reducing blood pressure, higher protein diets have been shown to improve insulin sensitivity and glycemic control (79, 81, 84, 94-96 ) while promoting greater weight loss (80, 83, 84, 97, 98) and improved long term sustained weight maintenance (99, 100) than low fat high carbohydrate calorie restricted diets. The weight loss superiority of higher protein, calorie restricted diets over either calorie restricted (low fat/ high carbohydrate) diets or calorie restricted (high fat/low carbohydrate) appears to be caused by the greater satiety value of protein compared to either fat or carbohydrate (97, 100-103). Of the three macronutrients (protein, fat, carbohydrate), protein causes the greatest release of a gut hormone (PYY) that reduces hunger (103) while simultaneously improving central nervous system sensitivity to leptin (97), another hormone that controls appetite and body weight regulation.
Dietary Protein and Bone Health
One of the crucial issues regulating bone mineral health and integrity is calcium balance which represents the difference between the amount of dietary calcium which is absorbed and the amount of calcium leaving the body through the urine and feces. Figure 5 demonstrates two key points: 1) most (~75%) of dietary calcium is not absorbed, and 2) calcium absorption increases with decreasing dietary intakes and decreases with increasing dietary intakes (104). Because dietary protein has been frequently, but not always (105-108), shown to increase urinary calcium excretion, it is possible that long term ingestion of high protein diets could lead to accelerated loss of calcium from the bones thereby impairing bone health and integrity.
Without the concurrent measurement of dietary calcium absorption along with urinary calcium losses the net calcium balance cannot be known. Hence, the simple observation that dietary protein ingestion may increase urinary calcium losses tells us little or nothing about calcium balance. In evaluating the effect of high protein diets upon bone mineral health, it is therefore crucial to measure both urinary calcium excretion and intestinal absorption of calcium. In this regard, Pannemans and colleagues (109) compared a low protein (12 % energy) to a high protein diet (21 % energy) in young and elderly subjects. Both a higher urinary calcium excretion and a higher intestinal absorption of calcium were induced by the high protein diet, thus no negative calcium balance occurred. A similar experiment confirmed that elevated dietary protein enhances calcium absorption and thereby counters the increased urinary excretion of calcium (110). Furthermore, a series of recent dietary interventions in humans has shown that high protein, meat based diets do not cause loss of calcium from the skeleton, but actually have a favorable effect upon it by lowering bone resorption (105, 107, 111, 112) and may actually increase bone formation by dietary protein induced increases in IGF-1 (105).
Figure 5. Relation between Calcium Intake and Absorption
Dietary Protein and Kidney Function
One of the most common misperceptions about high protein diets is that they can damage the kidneys of healthy normal individuals. This concept is known as the “Brenner Hypothesis” (113) and suggests that increased dietary protein elevates the kidney’s filtration rate (GFR) which in turn alters the kidney’s structure (glomerulosclerosis) which then causes albumin to appear in the urine (microalbuminuria). Although these series of steps represent the hypothesis Brenner proffered (113), his experiments actually showed an entirely different series of events. In reality, Brenner demonstrated that patients with pre-existing kidney disease had an elevated GFR, glomerulosclerosis and microalbuminuria and that by reducing dietary protein the GFR and microalbuminuria could be lowered (113). He further suggested that because elevated dietary protein increased the GFR in short term studies (< 2 weeks) of healthy normal subjects, protein was responsible for kidney damage. The problem with this interpretation is that markers of functional kidney damage in the normal subjects (microalbuminuria) were not demonstrated along with the elevations in GFR, nor were any long term studies (3-6 months) carried out to determine if the kidneys adapted to a higher protein intake.
The incidence of diabetic end stage kidney disease has increased steadily over the past three decades (114, 115). If dietary protein were responsible for causing kidney damage, then one might expect that dietary protein would have steadily increased during this same time interval. In fact, dietary protein significantly declined during this same period (19). Further, in the National Health and Nutrition Education Survey (NHANES III) of more than 12,000 subjects, dietary protein intake was unrelated to the incidence of microalbuminuria in normal, healthy subjects (117). In a comprehensive review of all randomized controlled trials evaluating the effectiveness of low protein diets upon GFR and kidney function, 11 studies (total n = 1223) showed no therapeutic effect, while only three investigations (total n = 235) demonstrated a significant benefit of dietary protein restriction on kidney failure progression (117).
The reason high protein diets do not have adverse effects upon kidney function in healthy, normal subjects may lie in the ability of the kidney to adapt to a higher protein intake. In a randomized controlled dietary intervention lasting 6 months, Skov and colleagues evaluated the effect of high (25 % energy) and low (12 % energy) protein diets upon kidney function in 65 healthy male and female subjects (118). The high protein diet did not cause urinary albumin to increase. Additionally, the specific GFR, which is an expression of the filtration rate per unit kidney volume, did not change during the high protein diet, indicating that renal (kidney) adaptation occurred to the higher protein load. The authors summarized, “We therefore conclude that a high dietary protein intake does not appear to have adverse effects on renal function in individuals without renal impairment.”
Dietary Protein and Cancer
Observational epidemiological studies frequently (119), but not always (120) show that high animal protein diets may increase the risk for a variety of cancers, particularly colorectal cancer (121). Consequently, it might be expected that non-meat eating vegetarians would have a lower risk for these cancers. Paradoxically, this effect has not been consistently demonstrated (119). A proposed mechanism of action for the carcinogenic effect of meat consumption is the formation of toxic N-nitroso compounds (NOC) in the gut from heme iron in meat (122, 123). Short term human studies are in agreement that increased meat consumption increases NOC formation both in the lower (122) and upper (123) gastrointestinal tract. However, whether this situation translates into increased cancer risk is not known because to date, no randomized controlled trials of increased meat consumption in humans, using cancer diagnosis as an end point, have been conducted.
The meats and fish consumed by pre-agricultural humans were almost always fresh, whereas current western diets contain significant quantities of processed, salted meats and fish preserved with nitrites and nitrates. Processed meats contains 10 times more NOC (5.5 µmol/kg) than fresh meat (0.5 µmol/kg) (124). Pre-agricultural humans consumed their fresh meats along with high intakes of fresh fruits and vegetables estimated to be between 35-45 % of total energy (14) compared to 8.1 % of total energy in the current U.S. diet (125). Increased fruit and vegetable consumption increases the fecal transit rate so that NOC have less contact time with the colonic mucosa and therefore may reduce the carcinogenic risk (126). Hence, the context under which high meat consumption occurred in hunter-gatherers varied significantly from what occurs in westernized populations. Animal based foods were almost always consumed fresh in conjunction with copious quantities of fresh fruits and vegetables. Even when vegetable intake was low or absent in these peoples, there is little evidence for an association of high protein, animal based diets with colorectal cancer. Prior to western acculturation, the Inuit may have consumed more than 95 % of their daily energy from animal and seafood (15), yet a comprehensive review examining virtually all historical and ethnographic data of these people prior to westernization was unable to document a single case of colorectal cancer (126). Should a high protein meat based diet initiate or promote colorectal cancer, then one might expect obligate carnivores such as cats to demonstrate high incidences of these malignancies. In, fact the opposite is true, and the rate of gastrointestinal tract cancers is quite low in domestic cats (128). In summary the case for animal based, high protein diets causing colorectal cancer, within the context of pre-agricultural diets, is weak.
Dietary Protein and Muscle Protein Synthesis and Fatigue
For athletes and individuals engaging in regular exercise, an animal based, high protein diet may be ergogenic and facilitate improved performance because of the stimulatory effect of dietary branch chain amino acids (BCCA) upon muscle protein synthesis (129-131), particularly when they are consumed in the post exercise window (132, 133). Table 2 demonstrates that lean meats and fish are much richer sources of the branch chain amino acids (valine, leucine and isoleucine) than are plant foods. In addition to facilitating muscle synthesis during the post exercise recovery period, BCCA may also improve endurance performance by reducing perceived exertion and mental fatigue by reducing the synthesis of brain 5-hydroxytryptamine, a substance that may promote central fatigue (134).
Table 2. Average branch chain amino acid (isoleucine, leucine and valine) concentration in various food groups (1000 kcal samples)
Dietary Protein: Summary and Conclusions
The evolutionary evidence indicates that so called “high protein diets” (20 – 30 % total energy) and “very high protein diets” (30- 40 % total energy) actually represent the norm which conditioned the present day human genome over more than 2 million years of evolutionary experience. The evolutionary template would predict that human health and well being will suffer when dietary intakes fall outside this range. Hence the current U.S. consumption of protein (15 % total energy) may not optimally promote health and well being. There is now a large body of experimental evidence increasingly demonstrating that a higher intake of lean animal protein reduces the risk for cardiovascular disease, hypertension, dyslipidemia, obesity, insulin resistance, and osteoporosis while not impairing kidney function.
How Much Protein Is Needed?
T. Colin Campbell, PhD
Jacob Gould Schurman Professor Emeritus Of Nutritional Biochemistry
Cornell University
Determining how much protein to consume is not a simple exercise. Protein, which is distinguished from its macronutrient carbohydrate and lipid partners by its nitrogen (N) content, is an essential nutrient. Philosophical, cultural and economic concerns have influenced the discussion on how much protein to consume ever since it was first discovered and named in 1839 by the Dutch chemist, Gerhard Mulder.
A good place to begin understanding how much protein to consume is with the recommended daily allowance (RDA, now recommended daily intake, RDI) that has been repeatedly re-evaluated and re-established for the past 60-plus years by many official bodies. Probably the simplest method of determining recommended intakes, at least conceptually, has been the N balance study. An experimental determination is made of how much protein, analyzed according to its N content, must be consumed in order to compensate for the average daily amount of N lost from the body.
After adjusting for the 16% N content of protein and after adjusting upward the experimental mean by two standard deviations to include the needs of about 98% of the larger population, this gives an ‘allowance’, or recommendation, of 0.8 gm of protein to be consumed per day per kg of body weight, for both sexes. This is equivalent to 48 gm of protein for a 60 kg adult female (132 lb), or 56 gm for a 70 kg (154 lb) adult male. Because this is based on the same dietary protein level for both sexes, I prefer these amounts to be expressed as percent of total diet calories. Along with the consumption of 0.8 gm of protein per kg/d, about 35 calories per kg/d is also consumed (i.e., 2450 calories for a 70 kg person). Therefore, 56 gm of protein per day represents 224 calories (1 gm of protein = 4 calories), or only 9.1% of the total 2450 calories consumed per day.
Said another way, a 9% protein diet theoretically is in excess of the amount of protein needed by about 98% of the population. Although some adjustments have been made for growth, pregnancy and lactation, a dietary protein content of about 10% is generally considered adequate for good health, for both sexes and for all ages.
The question, then, should not be how much protein is needed but how much protein in excess of the 10% dietary level can be safely consumed. This a highly relevant question because, according to national surveys, diets containing from 11% to 22% protein (mean of about 16%) are routinely consumed by about 90-95% of the American population. Said another way, the question is what health benefits and/or risks might be expected by consuming protein in excess of the amount needed?
At this point, it is worth noting that most people, for a very long time, have been concerned about getting enough protein. Many have also assumed that animal-based foods are the best source of protein because of their higher ‘quality’ or ‘biological value’, an underlying assumption of my own doctoral research dissertation about 45-50 years ago. The scientific community has indeed encouraged this public assumption ever since the discovery of protein. Mulder, for example, in 1839 gave this N-containing substance its name from the Greek word, proteios, which means ‘of prime importance’. Justis von Liebig, a contemporary of Mulder, went on to state that protein was the stuff of “life itself”. The famous German nutritional chemist Karl Voit and his many prominent students in the late 1800s and early 1900s, recommended that 100-130 gm per day should be consumed even though Voit, himself, found that 52 gm per day was sufficient for good health. Voit’s student, Max Rubner, also a prominent nutrition researcher, said that protein interchange was “the right of civilized man” while another Voit student, A.O. Atwater, went on to encourage similarly high protein intakes while he was founding the USDA laboratory in the U.S.
Importantly, these early nutrition scientists, ultimately becoming ‘fathers’ of their field, almost always equated protein with animal flesh. Although they acknowledged that plants also had protein, it was of “lower” quality, so they said, thus laying the groundwork for the idea that plant proteins had nutritional value only when they were carefully combined in order to compensate for their respective deficiencies of certain amino acids.
The personal biases of these early investigators often were expressed with considerable hostility and arrogance. The medical director of the British Empire Office in India, for example, offered that those who live on plant-based foods were of a “cringing and effeminate nature”.
In sharp contrast, Professor Russell Chittenden of Yale University School of Medicine, offered a very different set of findings, as published in two large books in 1904 and 1907. He showed that a group of young military recruits could obtain excellent physical conditioning over a 6 month period, as examined in 15 strength and endurance tests, by consuming a mostly plant protein-based diet. Not to be misunderstood that it might be the training and not the diet that led to these results, he then showed even greater conditioning when a group of already fit athletes switched to a plant-based diet. These remarkable and highly significant findings came at a time when Voit and his students and associates were pushing for higher, not lower, intakes of animal-based protein. Later, in 1922, the famed Dr. Benjamin Spock who was at that time a member of the Olympic crew team at Yale, was being advised along with his team mates to consume a high animal protein diet. He learned of Professor Chittenden’s work at his university only many years later when he wrote to me shortly before his passing and wondered why his coaches had made no mention of their colleague’s work. Little did Spock know that, about that same time in the early 1920s, Chittenden was being severely chastised for his work by his professional colleagues. Was he, perhaps, of an effeminate nature?
Let’s now return to the question of what happens when protein intakes are increased beyond 10% of calories, say from about 10% to 20% of calories, the range for most Americans. A variety of adverse health effects have been demonstrated and these effects are remarkably consistent among clinical, experimental (i.e., intervention), and observational type studies.
At the clinical level, for example, animal-based proteins (primarily) act on fundamental biological systems such as acid-base buffering, hormonal messaging, and enzyme activities that set the stage for a wide variety of adverse health outcomes. Thousands of studies have demonstrated these effects.
For example, the consumption of animal-based proteins can lead to a chronic condition known as metabolic acidosis, which has been known since the late 1800s. Secondary effects of this condition are extensive and include, for example, bone calcium loss to buffer the excess acid (thus weakening the bones) and inhibition of the critical enzyme that controls the formation of the ultimate vitamin D metabolite now known to be involved in a wide variety of health promoting effects.
Hormonal profile distortion, for example, includes elevated estrogenic hormone activities (higher estradiol and lower sex hormone binding globulin) and its corollary effects on breast cancer risk and other steroid responsive tissues. Also, animal protein consumption increases growth hormone activities like IGF-1 and IGF-2 that now associate with the promotion of multiple cancers.
Altered enzyme activities can be quite dramatic, occurring within hours after consumption of increased amounts of animal-based proteins and, further, these effects can be sustained for long periods of time. These dietary protein effects are especially notable for an enzyme complex involved in a broad array of reactions, including chemical carcinogen activation, steroid biosynthesis and disposition and fatty acid metabolism.
These clinical effects are consistent both with human intervention studies designed to control or even reverse serious chronic diseases and with human population studies. Increasing intakes of animal protein within the 10% to 20% range increases urinary calcium loss, elevates total and LDL cholesterol levels, increases cell replication and enhances DNA damage resulting from enzymatic activation of chemical carcinogens. These and many other clinical effects parallel higher incidence and/or mortality rates of osteoporosis, cardiovascular diseases, certain autoimmune diseases and various cancers, among other ailments. These findings are only a small sample of animal protein induced disorders and are cited here because of the consistency of evidence across clinical, experimental and observation type studies within the 10-20% range of dietary protein experienced by most Americans.
Although these outcomes and observations mostly refer to the independent effects of protein, it should be noted that the isolated effects of individual nutrients usually represent only a small part of a much larger story. First, the activities of nutrients studied in isolation are often quite different than when they are studied in the context of food. Second, when animal protein-based food consumption is increased, plant protein-based food consumption is usually decreased. Thus associations of animal-based foods with various disease outcomes almost invariably include simultaneous contributions resulting from the absence of the protective factors in plant-based foods. Plant-based foods have a wide variety of constituents that tend to oppose, or re-balance, the adverse effects of animal-based foods on these fundamentally important biological systems.
These are only a few of the effects of increasing total dietary protein above 10% of diet calories, mostly by the addition of animal protein-based foods. But these findings are cited here because (1) they show effects of animal-based protein on fundamental biological systems, (2) they are consistent across different types of studies and (3) they occur within the 10-20% dietary protein range. What makes this more generalized information especially compelling is its ability to explain a group of diverse yet remarkably profound diet and health observations reported during the past few decades. In population studies, for example, animal protein intake exhibits very impressive correlations (80-90%) with breast, prostate, ovarian, kidney and colon cancer, with increased disease risks appearing at very low levels of dietary animal protein (i.e., the regression line passes through the origin). Similar correlations also exist with cardiovascular diseases, type I and II diabetes, hypertension, osteoporosis, multiple sclerosis and kidney stones, among other ailments—all within the 10-20% dietary protein range.
Moreover, dramatic reversal, even cure, of advanced degenerative diseases as well as relatively non-fatal ailments, have been observed by switching away from animal protein-based foods to whole plant protein-based foods. The demonstrated reversal of advanced heart disease in humans is truly astounding, as is the 34 years-long control of multiple sclerosis progression. And finally, our own years-long series of experiments with laboratory rodents are especially informative of the fundamental effects of animal protein on experimental cancer development. When casein (cow’s milk protein), for example, is fed above the level that meets physiological needs, tumor growth progresses; when casein is fed at levels below 10%, tumor growth is suspended or reversed. That is, we could turn on and turn off experimental tumor development by feeding or withdrawing dietary casein at levels above or below 10% of calories, respectively.
I am convinced that many of the most cherished assumptions of nutritional science can trace their origin to our long and unquestioned reverence for protein, especially for animal-based protein. Diets have been constructed and judged according to their inclusion of this treasured nutrient. Diet and health policy has been structured in such a way so as to protect the consumption of animal-based protein. The first ‘official’ recommendation by the National Academy of Sciences in 1982 to decrease fat intake only to 30% of diet calories—even though the evidence at that time indicated lower fat intake was warranted—was to protect the consumption of animal protein-based foods. I know because, as a member of that committee, I wrote with a colleague the draft chapter on protein and cancer showing that animal protein intake was as much associated with cancer risk as was fat intake.
This long-standing reverence for animal-based protein often seems to be very personal, perhaps macho building, for some individuals. In point of fact, it may surprise many that when done right, consuming whole plant-based foods are known to lead to better not lesser athletic performance and physical fitness. Nonetheless, questioning the health value of animal-based protein still causes as much hostility today as before for some individuals, especially if they are economically or religiously conflicted.
My conclusion from these many observations is that animal protein, when added to diets already containing 10% protein, has the potential to promote the development of a wide variety of serious and oftentimes fatal diseases—not only because of the direct effects of protein but also because of the many parallel effects of companion nutrient imbalances created by animal-based foods simply to get access to the extra protein. I find that 10% total dietary protein, which has been long recommended by multiple policy committees and acknowledged in scientific reviews, is not only adequate but also is relatively devoid of risk of serious diseases and other ailments. It just so happens that 10% dietary protein is the same level that is typically found in a diet of varied whole plant-based foods. Increasing dietary protein above this 10% level generally means adding or substituting animal-based foods for plant-based foods but doing so invites a plethora of adverse health effects. To be very clear, I believe that total dietary protein should be 10% of calories, with virtually all of it being in the form of plant-based protein.
This commentary primarily addresses the effects of dietary animal protein because this was the mandate given to me. But I should emphasize that among the various adverse dietary effects associated with increasing consumption of animal protein-based foods, there is often the possibility that these effects are attributed, at least in part, to the consumption of highly processed foods rich in fat, salt and refined carbohydrates. In many studies, it is not easy to differentiate these separate effects, in part because of inadequate data. Nonetheless, prominent animal protein effects have been observed and mechanisms have been defined for the singular effects of this nutrient. In the final analysis, however, consuming a diet comprised of whole plant-based foods solves both problems caused by animal-based and highly processed foods. It is clear to me that a diet comprised of whole vegetables, fruits and cereal grains (unless otherwise contraindicated by allergic responses) with little or no animal-based foods, resulting in a dietary protein level of about 10%, is as near to an ideal diet as one can get.
Rebuttal to:
T. Colin Campbell, Ph.D.: How Much Protein Is Needed?
Loren Cordain, Ph.D., Professor
Department of Health and Exercise Science
Colorado State University
Rebuttal Overview
For the readers I think it is necessary to state that I have no personal enmity towards Colin – rather the opposite. I have met him at a nutritional conference in Boulder, Colorado a few years ago, and we had a warm and cordial conversation. My sole goal with my first paper and with this rebuttal is to seek the most accurate scientific truth that the available data can provide regarding the role dietary protein plays upon human health. Above all, the data must speak for itself without the intervening prejudice introduced by charismatic personalities, faulty human judgment and preconceived biases. I repeat: the data must speak for itself. Scientific theories must solely be driven by informed hypotheses based upon objective, well controlled experimental data. When the data consistently do not support the hypothesis, it is inappropriate to manipulate and selectively use flawed data (either knowingly or unknowingly) to continue to support the erroneous hypothesis (135). Initially, no matter how contrary they may appear to common sense, new hypotheses must be tested to replace previous hypotheses that no longer are consistent with the best available data (135).
I have organized my rebuttal into two categories: 1) general comments, and 2) specific comments. Within the general comments I will point out the generally flawed scientific logic that Colin follows to form his hypothesis that high protein diets have deleterious health effects. Within the specific comments, I will show you, point by point, how Colin has overstepped the bounds of the scientific method by misinterpreting data and selectively ignoring key studies that demonstrate the therapeutic effect of high protein diets.
In this rebuttal I have employed the reference numbering system I used in my opening paper, and any additional references I use in this rebuttal begin with the number, 135.
General Comments
The scientific study of human nutrition represents a sub-discipline within the much greater discipline of biology. Over the past 100 years, remarkable “discoveries” have been made into how dietary factors influence our health and well being. For instance, the “discovery” that vitamin B12 was an essential component of the human diet, without which our health suffers greatly, was made as recently as 1948 (1). The “discovery” that diets devoid of animal food cause blood and urine concentrations of the conditionally essential amino acid, taurine, to decline has only been known since 1988 (40).
A more accurate description of “discoveries” in biological sciences (in which the study of nutrition falls) is the uncovering of pre-existing phenomenon. The human requirement for vitamin B12 has been in place for eons before we uncovered what was already there. The human requirement for vitamin B12 and the decline in blood concentrations of taurine with reduced animal protein occur, not because scientists have “discovered” them, but rather because these fundamental mechanisms are woven into the fabric of our genes. Humans have a dietary requirement for vitamin C because we lack a functional gene that codes for the enzyme (L-Gulono- γ- lactone oxidase) which normally allows glucose to be converted to vitamin C. Analysis of the human gene coding for L-Gulono- γ- lactone oxidase shows that it has acquired a number of mutations (via natural selection) over the past 70 million years which have rendered it inactive (136). Because our primate ancestors ingested sufficient vitamin C containing foods, mutations which inactivated this gene had little negative influence upon survival and reproductive success of individuals bearing these genes.
Any unified theory of human nutrition represents a detective story in which scientists attempt to reveal or uncover physiological systems that have been designed by, and put in place by evolution through natural selection. Accordingly, hypotheses regarding what modern day humans should and shouldn’t eat must be consistent with the system and the ancient environmental selective pressures that engineered our current genes. If we are to buy into Colin’s hypothesis that optimal human health occurs when dietary protein represents about 10% of energy, then the system (evolution via natural selection) which shaped our present genome necessarily had to be conditioned over eons by a low protein intake. Otherwise, there is no rational alternative hypothesis to explain why humans have a low dietary protein requirement. A low dietary protein requirement for optimal human health can only occur if it is dictated by our present day genes which in turn are shaped by events that have occurred in the environment of our ancestors in the distant and remote past.
As I have laboriously and meticulously lain out in my initial essay, there is no credible fossil, archeological, anthropological, anatomical, ethnographic or biochemical evidence to show that members of our genus (Homo) routinely consumed low protein diets. In fact, without the inclusion of energetically dense animal food into the hominin diet, starting at least 2.5 million years ago, our large energetically active brains would not have evolved (16, 34, 35). Accordingly, the fundamental logic underlying Colin’s hypothesis (that low protein diets improve human health) is untenable and inconsistent with the evolution of our own species.
Specific Comments
1. Page 18, column 1, paragraph 1. As Colin has suggested, nitrogen balance studies have been extensively used to predict the minimal human protein requirements (137). Because all protein contains 16.6% nitrogen, then the measurement of nitrogen serves as a proxy for protein. Colin’s argument is that optimal health occurs when we are in zero nitrogen balance (e.g. when our daily nitrogen intake = daily nitrogen excretion), and that excessive protein intake and hence negative nitrogen balance (nitrogen intake < nitrogen excretion) causes ill health and disease. Colin suggests that zero nitrogen balance occurs when the daily protein intake is 10 % of energy.
The problem with this oversimplification is that minimal protein requirements provide no information about optimal protein requirements. As is the case with many nutrients, the minimal protein requirement may not necessarily represent the optimal requirement. The minimal vitamin D requirement is 600 IU, however additional therapeutic effects occur when the daily intake is considerably greater (138, 139). Similarly, niacin (vitamin B3) is of therapeutic benefit in lowering blood cholesterol concentrations at daily intakes much higher than daily recommended intakes (DRI) (140). As I have extensively outlined in my first paper, a large body of experimental evidence now demonstrates a higher intake of lean animal protein reduces the risk for gout (49), cardiovascular disease (70, 74-86), hypertension (85, 87-93), dyslipidemia (74-86), obesity (80, 83, 84, 97-103), insulin resistance (79, 81, 84, 94-96 ), and osteoporosis (105, 107, 109-112 ) while not impairing kidney function (116-118).
2. Page 18, column 2, paragraph 4. It is difficult to objectively analyze experiments conducted more than 100 years ago in regard to their present day validity (141). As Colin has indicated, the design of this experiment was such that the exercise training effect could not be separated from the dietary effect. Hence the improvement in strength and endurance shown in this experiment cannot necessarily be attributed to the plant based diet, particularly when more recent, well controlled studies have shown otherwise.
As I have previously pointed out in my original essay, plant based diets contain much lower concentrations of the ergogenic branch chain amino acids (BCAA) (valine, leucine and isoleucine). For instance, a 1000 kcal serving of lean meat contains on average 33.7 grams of BCAA whereas a comparable serving of whole grains contain a paltry 6.1 grams of BCAA. High protein diets are ergogenic because of the stimulatory effect of dietary BCCA upon muscle protein synthesis (129-131), particularly when they are consumed in the post exercise window (132, 133). Additionally, BCCA may also improve endurance performance by reducing perceived exertion and mental fatigue by reducing the synthesis of brain 5-hydroxytryptamine, a substance that may promote central fatigue (134).
3. Page 19, column 1, paragraph 3. In regard to the metabolic acidosis produced by meats and their potential for adverse health effects, it should also be pointed out that cereal grains (a foundation of plant based diets) maintain an acid load similar to meat and animal foods (142). Whereas lean animal protein consumption has been shown to have a favorable effect upon bone mineral health by lowering bone resorption (105, 107, 111, 112) and increasing formation (105), whole grain cereal consumption impairs bone mineral health (143). In populations where cereal grains provide the major source of calories, osteomalacia, rickets and osteoporosis are common place (144-147). In animal studies it has been long recognized that excessive consumption of cereal grains can induce vitamin D deficiencies and bone mineral abnormalities in a wide variety of animals (148-150) including primates (151). The mechanism by which cereal grain consumption promotes bone mineral disorders may be via their ability to interfere with the entero-hepatic circulation of vitamin D or its metabolites (151, 152), or by increasing the rate of inactivation of vitamin D in the liver (153).
4. Page 19, column 1, paragraph 6. As I have previously pointed out, increased loss of urinary calcium by high protein animal based diets does not necessarily result in skeletal calcium loss (105, 107, 111, 112 ) because calcium absorption is simultaneously increased (109, 110). In regard to lean animal protein elevating total and LDL cholesterol, I am not aware of a single study which has demonstrated this effect, particularly when the effect of saturated fat has been eliminated. In fact, the opposite is true. Increasingly, experimental evidence in humans shows that lean animal protein improves all facets of the blood lipid profile (79-85 ).
5. Page 19, column 2, paragraph 2. As I have previously pointed out in my initial essay, observational epidemiological studies cannot show causality (68, 69). In addition, not all epidemiological studies show strong or “impressive” relationships between animal protein consumption and various cancers (120, 154, 155), particularly when the effect of high cooking temperatures is eliminated (156). Observational epidemiological studies showing “correlations” to type 2 diabetes, hypertension, osteoporosis and kidney stones fall by the wayside and become meaningless when more powerful human dietary interventions show that high protein diets are actually protective for these diseases and their symptoms, as I have extensively reported in my first essay.
6. Page 19, column 2, paragraph 3. In this paragraph Colin suggests that reversal of a number of degenerative diseases can occur when animal protein based foods are replaced with whole plant protein based foods. Unfortunately, no references are provided so it is difficult to comment or rebut these remarks. However, I can comment upon the extensive rodent experiments that were conducted in Colin’s laboratory over more than a decade in which he examined the role of milk protein (casein) in the development of liver cancer.
Aflatoxins are naturally occurring toxins that are produced by a variety of fungi, most notably Aspergillus flavus and Aspergillus parasiticus. These fungi normally reside in soil, but can invade food crops under conditions of high humidity or other stress. Food crops most frequently affected are cereals, nuts and legumes (which represent the mainstays of plant based, vegetarian diets). The toxin is also found in the milk of animals fed contaminated feed. Aflatoxins are metabolized in the liver to become potent liver carcinogens for all mammals including humans (157). High-level aflatoxin exposure causes liver cancer, whereas chronic, low level exposure to aflatoxin does not necessarily lead to cancer (157).
Colin’s research group developed a rodent model of liver cancer in which they dosed the animals with high concentrations (typically 200-350 microgram/kg per day for 10 days) of aflatoxin and then fed them diets containing varying amounts (typically 20, 16, 12, 8, 4 % of total energy) of casein (158-161). Regardless of the casein dose, all animals developed cancerous or pre-cancerous liver lesions (161), however the animals fed the higher amounts of casein developed more cancerous lesions, particularly when a level of approximately 12 % casein was reached (160). From these experiments Colin concluded that, “low protein intake inhibits lesion development” and that “AFB1 (aflatoxin) induced preneoplastic foci (precancerous lesions) depends upon a high protein intake” (161).
Although Colin has inferred from his experiments with rodents that high protein diets promote cancer and low protein diets repress it following cancer initiation by a carcinogen, this interpretation is incorrect. The only logical conclusion that can be reached from his series of experiments is that only the milk protein, casein, when consumed at more than 10% of energy, promotes liver cancer in rodents exposed to high concentrations of aflatoxin. His experiments cannot be generalized to other animal proteins, such as those found in lean meats. In the typical U.S. diet, dairy food consumption represents 10.6 % of the total energy (13) and is distributed in the following manner: whole milk (1.6 %), low fat milk (2.1%), cheese (3.2 %), butter (1.1 %), and other 2.6 % (13). The total protein content by energy for milk is 21 % , low fat milk ~29 % , cheese ~25 % , butter 0 %, and other ~27 %. Casein represents ~80 % of all milk proteins, so the average total casein content in the U.S diet is ~2.0 % energy. Accordingly, current consumption of casein in the U.S. diet would have little or no bearing on cancer incidence rates if we assume Colin’s rodent model of cancer is correct and applicable to humans.
In the typical U.S. diet ~75 % of the protein comes from animal products other than dairy (20). In the U.S. diet muscle meats from birds, mammals, fish and invertebrates represent far and away the greatest protein source, as organ meats are infrequently consumed. The primary proteins in muscle tissues are actin and myosin. Consequently, Colin’s rodent experiments using casein as a generalized surrogate for protein in the typical U.S. diet has little or no relevance to human cancers for two reasons. First, as I have previously shown, in the U.S. population it would be virtually impossible to ingest 10 % of the daily caloric intake as casein, and secondly the primary animal proteins (actin and myosin) in the typical U.S. diet were never tested in Colin’s animal experiments. Hence Colin’s conclusions that high dietary protein from all animal sources promotes cancer and lower dietary protein prevents it cannot be inferred from his rodent experiments.
7. Page 20, column 2, paragraph. Colin’s statement “It is clear to me that a diet comprised of whole vegetables, fruits and cereal grains (unless otherwise contraindicated by allergic responses) with little or no animal based foods, resulting in a dietary protein level of about 10%, is as near to an ideal diet as one can get” clearly reflects the bias he brings into this debate. If only Colin would have framed his hypothesis from an evolutionary perspective years ago, he would have realized that his viewpoint of an ideal diet is erroneous and untenable.
It may surprise Colin, but cereal grains were rarely or never consumed by all of humanity until very recent times (< 500 human generations) (13, 143). Additionally, the incidence of celiac disease (caused by consumption of wheat) is now estimated to afflict 1 in 133 people in the U.S. (162). What should the ideal diet be for these 2,255,639 U.S citizens? And why might autoimmune diseases be caused by grain consumption in the first place (143, 163)? Finally, if vegetarian diets are “as near to an ideal diet as one can get” why then do meta-analyses of all cause mortality (including cancers) in vegetarians show them to be no better off than the general public? (164).
Rebuttal to:
Loren Cordain, PhD: The Evolutionary Basis for the Therapeutic Effects of High Protein Diets
T. Colin Campbell, PhD
Jacob Gould Schurman Professor Emeritus of Nutritional Biochemistry
Cornell University
My critique of Professor Loren Cordain’s proposition almost entirely depends on my philosophy of nutrition. It is clearly different from that of Cordain’s understanding of this discipline. He mocks the science of nutrition as if it has little or nothing to offer. I believe that it has much to offer even though its essence mostly remains hidden. He says that nutritional science is a “newly established discipline”, that it “is a highly fractionated, contentious field with constantly changing viewpoints”, that it “remains an immature science” and “that there is such seeming chaos, disagreement and confusion”. He then suggests “nothing in nutrition seems to make sense because most nutritionists have little or no formal training in evolutionary theory”. He apparently believes that only paleontologists and archeologists have a crystal ball as to what is nutrition and, more specifically, how much protein should be consumed.
He has thrown down a challenging gauntlet, thus leaving me no option but to explain what I believe nutrition to be—before I tackle the protein requirement question.
While I agree that there is enormous confusion, both in the professional and lay communities, I disagree that nutrition is a relatively “new” science and that hypotheses about nutrition must pass a test set by archeologists before they can be considered reliable. I agree that nutrition hypotheses should be consistent with our evolutionary past, but having to pass an archeologist’s ‘smell’ test is too restrictive based on the evidence presented in Cordain’s paper.
Protein was discovered in 1839, fat and energy were being discussed in the mid 1800s and amino acids were known to be components of protein in the late 1800s. I suggest that nutrition as a science predates the well-known disciplines of genetics, bacteriology, virology, biochemistry, pharmacology, immunology and molecular biology, to name only a few. Is it possible that Cordain considers nutrition to be a “new” science because it is new to him? Is it so confusing to him because he knows so little about the science?
Being confused about the concept of nutrition is a serious matter. It is real—both inside and outside of the field. Many factors contribute to this confusion. In addition to explanations like ignorance, economic pressures and food preference biases, I believe that there is a more fundamental issue that underlies each of these explanations, this being the way we think about scientific investigation itself. More specifically, it concerns the role that precision, as a concept, plays in research. Ever since the Renaissance, scientific investigation has emphasized this concept when gathering empirical evidence. Physical measurements must be precise, substances and events must be precisely characterized physically and chemically, and hypotheses have more value if they are precisely focused (especially if one expects to compete for research funding). Even to question this concept of precision is scientific heresy to many scientists.
Making use of precise measurements is essential for the research and development (R&D) of complex physical technologies like automobiles, space shuttles, telephones, and computers. When developing these complex physical products, we precisely make the component parts then precisely assemble them into complex products. We must be mindful of precision and I am delighted for that.
But the same cannot be said for biomedical R&D, even though most researchers and other observers seem to believe otherwise. Although we may precisely know and control the components and activities of a biological event (as with a specific biochemical reaction), this is far, far removed from trying to ‘do’ R&D on complex products that we call life, disease or health. This is also true for ‘simpler’ products like body organs or even sub-cellular organelles.
Of course, there are times when precision in biomedical research is exceptionally important. For example, we can surgically penetrate minute bits of tissue, we can determine the exact chemical structure of each of the 30,000 or so human genes and we can precisely synthesize chemicals (i.e., drugs) that interact with these genes and their protein products. These are fascinating research journeys and sometimes they can produce real life benefits. But no matter how precisely measured and described are these events and their component parts, we cannot assemble them into complex biological outcomes like those fashioned over millions of years by Nature. We cannot construct biological complexity that is faithful to the natural order of things, no matter how precisely we measure and know the component parts of these complex systems. This is because each event or bit of matter of these systems is usually investigated in relative isolation from its native biological environment. In biological systems, concentrations and activities of the component parts do not exist in discreet quantities (as in physical systems) because they operate, in time, over a continuum. And making matters decidedly more complicated, countless higher order interactions among the component parts cannot be accurately described or controlled. When investigating the nature of complex intact biological systems, we cannot construct them de novo as in physical systems. We can only provide resources then simply observe the outcomes that are produced.
In brief, the criteria and considerations for doing research and development on physical systems are very different from doing research on biological systems. With physical systems, we are in control when doing research developing products. Measuring and assembling things with precision is in our best interest. We know the dimensions, the structural identities and the time parameters and we can stabilize them to our specifications. Except in the most narrowly defined systems, we cannot use precision to construct or even to accurately describe the far more complex and dynamic biological systems. Measuring things with precision does not have the same meaning in biology because the system is always changing in exceptionally complex ways. Moreover, the interactions between the component parts are also changing.
By exaggerating the importance of precision in biomedical research, we also err in other ways. That is, we too often misinterpret the concept of precision as perfection. Assuming more perfection implies more ‘value’ and for some scientists, precision may mean invincibility and absolute truth (or might we say ‘arrogance’?). A major impetus for this view of precision, cum perfection cum absolute truth, is the marketplace mentality where health products and information are valued more for their monetary value than for their health value. Monetary value can be measured precisely but, more importantly, intellectual property protection is enhanced with more precise knowledge.
Biomedical and nutritional research should be conducted within a paradigm that is substantially different from the contemporary, traditional paradigm. We must strive to develop hypotheses, to organize research studies, and to interpret and apply results that acknowledge biological complexity and that describe cause-effect relationships cognizant of that complexity. From this perspective, observations on the component parts, however precisely they may be measured, only represent relative truths, not absolute truths. Moreover, we should stress research that is observational, comprehensive and representative of real life conditions.
This same biological complexity describes the essence of nutritional effects or, perhaps, describes what nutritional science should be. Most serious students of nutrition know well this complexity even though many scientists ignore its implications.
The implications of this view are wide-ranging and probably more significant than most people realize. This view is ‘holistic’ and, among other considerations, means that deciding which groups of foods to consume is far more important than deciding what levels of individual nutrients should be consumed. It means that priority should be given to the search for collective disease and health outcomes that may respond to the same diet and lifestyle conditions. It means investigating a variety of metabolic events that mutually support maintaining homeostasis and regulating adaptive responses, then exploring whether there are groups of food to support this group of metabolic events. It means rejecting randomized clinical trials as the acclaimed ‘gold standard’ of diet and health research. It means questioning the validity of recommended intakes/allowances (e.g., RDAs) on a nutrient-by-nutrient basis. It means acknowledging that the functional activity of a nutrient within a food environment can be far greater than an equal chemical amount in a tablet. Most importantly, it means using ‘weight of evidence’—all evidence—as the standard method of determining the reliability of cause-effect hypotheses.
This is my philosophy, both of nutrition and of the scientific method itself. More than any other consideration, this philosophy makes it difficult to comment on a number of Cordain’s assumptions and observations, even if they may be accurately cited. I challenge many of Cordain’s assertions but, in the interest of space, I will comment only on a few of these points in the hope that an explanation of my philosophy will make clear my answers to the others.
Specific Comments
1. Figure 4, page 9. The randomized clinical trial (RCT) is NOT the “trump card” of all diet and health studies, as claimed. RCTs were meant for the testing of pharmaceutical agents and would-be pharmaceuticals called neutriceuticals (i.e., nutrients), not for assessing lifestyle nutrition. RCTs have undoubtedly distorted the concept of nutrition and caused more confusion in diet and health research than almost any other scientific endeavor in recent years.
2. Page 4, second column, last paragraph. The statement that “all human requirements… are ultimately genetically determined” is an oversimplification. Several studies, including a paper of our own over 25 years ago (Science 211:719-721), have shown that in utero exposure to an appropriate agent can cause a non-genetic (i.e., epigenetic) lifetime change that undoubtedly affects nutrient requirements. This finding, now reported in more recent studies by others, would have been incorrectly considered genetic.
3. Page 5, column 1, paragraph 2. The statement is made that “the upper limit of protein ingestion is determined by the liver’s ability to synthesize urea.” If this is an inference that all could be well until this limit is reached, it is grossly misleading. According to Cordain, this equates to an astounding intake of about 246-270 g protein/day, or about 40% of total calories, 4-5 times the long-standing recommendation/allowance! I have 2 problems with this statement.
An adverse nutritional effect almost never results in a single outcome. Such effects occur serially over time, beginning with the least problematic and proceeding to the most problematic. Saturation of the urea disposition cycle is likely very serious but it will be preceded by many earlier adverse effects, some of which may take years to develop. Unfortunately, Cordain glossed over these effects as if they were unimportant. I refer, of course, to the numerous effects that I cited in my paper and that also are present in our book and in my many publications. I am suspecting that he believes that his archeological evidence is sufficient to make his main points and that modern day investigation of existing problems is nutrition gobbly-d-gook.
4. Page 4 and throughout. Cordain laments nutrition researchers as not being schooled in evolutionary theory. Because I was not trained in this discipline, I shall turn for advice to two professionals who were so trained. The first is a creative evolutionary psychologist (PhD, University of Virginia, Instructor at Stanford University), Dr. Doug Lisle, who with his colleague, Dr. Alan Goldhamer, wrote a fascinating small book, “The Pleasure Trap”. As you can see, Lisle also relies on some brilliant work of others (Dawkins and Williams) in this field. Lisle and I have talked about the basis for this high protein recommendation by anthropologists like Loren Cordain and he puts it this way:
“There is an important misconception among those who attempt to use evolutionary theory to guide their thinking in questions about human nutrition: that the diet consumed in human evolutionary history is the diet that optimally serves human health. This is a serious error, and one that makes major mistakes quite possible, even likely.”
Those who follow evolutionary history and who support the inclusion of substantial amounts of animal protein as a recommended dietary practice, make this error. Our dietary evolutionary history, while interesting, absolutely does not yield critical clues for optimal nutritional practices. Human evolution required that our ancestors make dietary choices that maximized gene proliferation. And that is absolutely the ONLY thing that such practices yielded (see Dawkins’, The Extended Phenotype, for clarity on these issues, also G. Williams’ classic Adaptation and Natural Selection).
“The reader may find an example to be helpful. Consider a dietary contest. Suppose that Diet A (a diet nearly exclusively of animal origin) resulted in 100% of individuals living long enough to reproduce, but despite adequate calories available throughout the lifespan, average non-accident/non-violent age at death (thus death was from nutrition-related diseases) was 50 years. Diet B (a diet exclusively of plant origin) resulted in just 60% of individuals living long enough to reproduce (perhaps because they were too finicky to eat meat [or because inadequate calories killed young people]), but if fortunate enough to be presented with an environment of sufficient calories (thus likely avoiding all nutrition related diseases)... life expectancy was 90 years. In this contest, Diet A wins the evolutionary battle, and is adopted. The evidence presented by modern nutritional science suggests that something akin to this may well be the case for humans, who have a large constellation of disease processes that can be laid at the door of animal foods, and effectively reversed with vegan diets (see Ornish, Esselstyn, McDougall, etc.). Seen from this perspective, speculation about human dietary history is a fascinating subject, but is actually irrelevant in the search for optimal nutritional practices. Such practices can only be determined by scientific investigation done in the present time, studying varying dietary practices and determining their comparative outcomes.”
I concur with this view, especially with the inference that present-day conditions need to be studied for present-day dietary advice, including the amount of protein to be consumed. Ample evidence now exists to show that our bodies are continually drawing on diverse non-genetic (i.e., epigenetic) adaptive mechanisms to enhance health and survivability even when the available resources are less than ideal. We adapt to inappropriate levels of nutrient intakes in ways that attenuate the adverse effects otherwise expected. And we do so without assuming that this is accounted for by genetic mutations, as suggested by Cordain (pp. 10-12). This does not, however, mean that we can fully accommodate inappropriate food without harm, only that these adverse effects are less than what they might otherwise be.
Cordain seems to believe that some of the most important evidence favoring high protein diets may be seen by considering the diets of present day hunter-gatherer societies that presumably reflect our evolutionary past. He uses an ethnographic atlas of 229 such societies and finds that “73% of the world’s hunter-gatherers obtained >50 percent of their subsistence from hunted and fished animal foods, whereas only 14 percent of worldwide hunter gatherers obtained >50 percent of their subsistence from gathered plant foods.” These appear to be impressive findings until one reads a rather devastating rebuttal of his findings that was published elsewhere by Professor Katherine Milton at the University of California at Berkeley (Am. J. Clin. Nutr. 71: 665-667). She first points out (1) that, on several grounds, this atlas is not a reliable compilation of dietary information, (2) that, in any event, another author using the same atlas concluded that “gathered vegetable foods were the primary source of subsistence for most of the hunter-gatherer societies he examined”, (3) that some of these “societies are largely free of diseases of civilization regardless of whether a high percentage of dietary energy is supplied by wild animal foods, wild plant foods, or domesticated plant foods taken from a single cultivar”, (4) that non-genetic adaptations are much more important than genetic mutations in determining diet responses, (5) that the “evolutionary history of anthropoid primates” going back 25 million years “shaped their nutrient requirements and digestive physiology well before they were humans or protohumans” (instead of Cordain’s suggestion that it was the last 2.5 million year history that shaped our nutrient requirements), (6) that “there is general agreement that the ancestral line (Hominoidea) giving rise to humans was strongly herbivorous”, being largely based on the consumption of tropical plant foods and (7) that the archeological evidence does not “shed light” on the proportionate consumption levels of animal and plant based foods. These seem to be powerful counter arguments to Cordain’s hypothesis.
Again, our own research supports these views. When metabolic control systems are compared across species as to their responsiveness to food sources and as to their associations with health/disease outcomes, there is overwhelming support for a linkage between the consumption of whole plant based foods, health maintenance, and disease prevention (this diet, not so coincidently, contains about 10% of the total energy as protein). The fact that these systems go deep into our evolutionary past, far more so than even the 25 million year benchmark suggested by Milton, suggests that much of our nutrient requirements could have been fashioned very early in our evolutionary history (certainly more than 2-3 million years ago proposed by Cordain).
Regardless of which side of the issue one favors, I can’t help but wonder whether the discordance Cordain finds in the field of nutrition does not also apply to his own field of evolutionary history. Therefore, why must studies of contemporary diet and health associations by nutrition researchers be submitted for validation by evolutionary history buffs, as suggested by Cordain? Why not the other way round?
5. Page 9, column 1, paragraph 2. I regret to say that Cordain’s commentary on establishing “cause and effect between diet and disease” (p. 18) is markedly different from my views. Aside from my earlier comments on the virtually worthless value for randomized clinical trials for nutrition studies, I also do not agree with his views on cross-sectional, observational studies, as we conducted in rural China.
Traditionally, epidemiologists and others will claim, as does Cordain, that cross-sectional population studies (also called ecological and correlation studies), which compare population characteristics, cannot only be used to generate hypotheses about possible cause-effect variables, not to prove causation. I absolutely agree with this view when one assumes highly reductionist hypotheses. In fact, virtually all epidemiologists and biostatisticians also agree. However, the reductionist philosophy that allows this criticism does not apply in most instances of diet and health associations. In contrast, if one assumes a more holistic philosophy when considering diet-health causality—as one should—then this precaution about the worthiness of cross sectional studies is far too restrictive.
The holistic philosophy may be applied in research studies in at least two ways. First, one can comprehensively record many factors that may influence outcome (even many possible health and disease outcomes) in a search for consistent and biologically plausible causal factors. For example, we can determine the proportion of plant-based food factors (both exposure and metabolic) that are consistently associated in the same study with a particular outcome or group of outcomes. Second, we can interpret the results of such studies in the context of other studies, especially those that show biological plausibility. Many years ago (1991) I explained this view in the 896-page monograph of the China Project, then restated it in our trade book, “The China Study”. It’s unfortunate that Cordain uses old logic to criticize findings like ours without reading the rationale for that study.
6. Page 6, column 1, paragraph 3. I do not subscribe to the tenor and substance of this section suggesting such importance for genes and gene mutations in the evolution of the dietary influence on health. I already mentioned the epigenetic evidence for long-term evolutionary change that does not depend on genetic mutation. In addition, there also are extensive data now existing to show that it is not the presence or absence of genes, good or bad, that actually determines health outcomes. We now know that it is the nutritional control of gene expression that matters far more. In our own work on experimental animals, we were able to demonstrate a remarkable ability of nutrition to control gene(s) responsible for development of experimental cancers—this is briefly summarized in our book, “The China Study”. I should also note that it was the protein of cow’s milk (casein) that we used to show this remarkable nutritional effect on gene control, thus leading to our seriously questioning cow’s milk and its products on many grounds as a serious health food—a point that Cordain and I share.
7. Page 7, column 2, paragraph 1. Cordain cites an estimate of 4-8.5% of total energy as animal food for chimpanzees, our nearest living non-human primate, more or less agreeing with his suggestion of 3-5% (mostly insects, etc.) in a presentation of his at Boulder, Colorado, a couple years ago. In either case, he notes that this proportion was established sometime before 2.6 million years ago prior to the introduction of stone tool technology, inferring that this estimate may not be relevant for present day human consideration because of the greatly increased intake of animal foods up to at least 50% of total energy intake during Paleolithic times. If the ‘blueprints’ for nutrient intakes were established before 2.6 MYA, then why are these estimates of 3-8.5% (3-5%) irrelevant?
8. Page 11, column 2, paragraph 2. Relying on the hypothesized effects of N-nitroso compounds on gut tissue as an explanation of large bowel cancer is too narrow a view. It ignores factors such as bile acid secretion, increased anaerobiosis, dietary fat, dietary protein and an array of antioxidants as important modifiers. The claim that vegetarians do not have lower cancer rates is an overstatement; many studies show that cancer rates are somewhat lower. But more to the point, about 90% of vegetarians still consume dairy products, among other animal based foods, thus minimizing the difference in nutritional composition from non-vegetarian diets. It is somewhat surprising to me that we even observed statistically significant deceases in disease rates with such small differences in nutrient composition.
Summary
Although there are many more statements in Cordain’s paper that I do not support, I hope that these comments are sufficient. As stated in the beginning, my philosophy of nutrition is substantially different from Professor Cordain if, in fact, he even has a philosophy of nutrition, given his very negative views of this field. I admit that my views represent a paradigm shift from the traditional views of nutrition, including many of my colleagues in the field, and it is this very different world view that prompts so much of my skepticism of Cordain’s observations and views.
My paradigm of nutrition assumes that a holistic food effect is far greater than a reductionist nutrient effect. Not only does this view shape my interpretation of research findings, it also challenges the design of experimental studies, the assessment of food and health policy, the very nature of science itself, and the choices of what to eat.
Briefly, I strongly disagree with Cordain’s proposal that 15% dietary protein is likely to be too low. I say that is likely to be too high, especially if animal based foods are required to reach this level. My recommendation is about 8-12% total protein, with all or virtually all of it being plant-based. With whole plant-based foods, it is not easy to go above this range without adding animal based foods or over consuming high protein plant-based foods like legumes.
I arrive at this estimate of 8-12% total protein first by considering a wide variety of research evidence. I then use this evidence to decide what the total diet should be for good health, not only based on the hypothetically isolated effects of protein but also because of the combined and highly integrated effects of all the chemical compounds of this food. This is a diet comprised of a variety of nutrient-rich plant-based whole foods, with very little or no added fat, sugar and salt. I then ask what is the likely protein content of this diet and this is generally in the range of 8-12%. I then find it quite remarkable that so many of the adverse effects of diets higher in protein begin to show their effects at about 10-12% total protein, as summarized in our book, “The China Study”. I also find it quite remarkable that for many years multiple panels who have considered protein requirements arrive at a level of about 6-8% being the protein ‘requirement’ to match nitrogen loss, and about 8-10% being the allowance to hypothetically insure protein adequacy for 98% of the population.
These are my main broad based concerns with Cordain’s observations and conclusions.
1. I don’t find his evolutionary theory arguments to be persuasive. First, researchers in his own field cite considerable evidence to the contrary to refute his arguments concerning the contemporary hunter-gatherer societies. Second, the evidence presented by Cordain and others does not establish the proportion of energy provided by animal based foods. Third, I believe that he has put far too much weight on gene mutation events to describe the evolutionary basis for a normal, healthy diet and has overlooked the highly significant roles of epigenetic and adaptive mechanisms to also account for these same effects. This suggests that diet-disease associations observed in contemporary times are far more meaningful than what might have occurred during evolutionary times—at least since the last 2.5 million years or so.
2. His reliance on reductionist research findings and theory is at odds with the nutritional effect as it exists in reality. This is a serious flaw.
3. He omits the overwhelming findings on the adverse health effects of dietary protein as it increases within the range of 10-20% total protein. These increases are, in reality, almost always provided by increased consumption of animal-based foods.
4. Although Professor Cordain likely will not know this, he and others should nonetheless know of the remarkable healing effects now being routinely accomplished by my clinician colleagues as well as told to me by those who change to a low protein, low fat, whole foods plant-based diet.
References
1. Bogert LJ, Briggs GM, Calloway DH. Nutrition and Physical Fitness, Ninth Edition. W.B. Sauders Company, Philadelphia, 1973.
2. Dyerberg J, Bang HO. A hypothesis on the development of acute myocardial infarction in Greenlanders. Scand J Clin Lab Invest Suppl. 1982;161:7-13.
3. Jenkins DJ, Wolever TM, Taylor RH, Barker H, Fielden H, Baldwin JM, Bowling AC, Newman HC, Jenkins AL, Goff DV. Glycemic index of foods: a physiological basis for carbohydrate exchange. Am J Clin Nutr. 1981 Mar;34(3):362-6.
4. No authors listed. Position paper on trans fatty acids. ASCN/AIN Task Force on Trans Fatty Acids. American Society for Clinical Nutrition and American Institute of Nutrition. Am J Clin Nutr. 1996 May;63(5):663-70.
5. National Academy of Sciences, Institute of Medicine. Letter Report on Dietary Reference Intakes for Trans Fatty Acids, 2002. http://www.iom.edu/CMS/5410.aspx.
6. http://www.mypyramid.gov/
7. Willett WC, Stampfer MJ. Rebuilding the food pyramid. Sci Am. 2003 Jan;288(1):64-71
8. Krauss RM, Eckel RH, Howard B, et al. Dietary Guidelines: revision 2000: A statement for healthcare professionals from the Nutrition Committee of the American Heart Association. Circulation. 2000 Oct 31;102(18):2284-99.
9. Blanck HM, Gillespie C, Serdula MK, Khan LK, Galusk DA, Ainsworth BE .Use of low-carbohydrate, high-protein diets among americans: correlates, duration, and weight loss. MedGenMed. 2006 Apr 5;8(2):5.
10. Ornish D. Dr. Dean Ornish’s Program for Reversing Heart Disease: The Only System Scientifically Proven to Reverse Heart Disease Without Drugs or Surgery. New York : Random House, 1990.
11. Nesse RM, Stearns SC, Omenn GS. Medicine needs evolution. Science 2006;311:1071.
12. Dobzhansky T. Am Biol Teacher. 1973 March; 35:125-129.
13. Cordain L, Eaton SB, Sebastian A, Mann N, Lindeberg S, Watkins BA, O’Keefe JH, Brand-Miller .Origins and evolution of the Western diet: health implications for the 21st century. Am J Clin Nutr. 2005 Feb;81(2):341-54.
14. Cordain L, Miller JB, Eaton SB, Mann N, Holt SH, Speth JD. Plant-animal subsistence ratios and macronutrient energy estimations in worldwide hunter-gatherer diets. Am J Clin Nutr. 2000 Mar;71(3):682-92.
15. Cordain L, Eaton SB, Brand Miller J, Mann N, Hill K. The paradoxical nature of hunter-gatherer diets: Meat based, yet non-atherogenic. Eur J Clin Nutr 2002; 56 (suppl 1):S42-S52.
16. Cordain L, Watkins BA, Mann NJ. Fatty acid composition and energy density of foods available to African hominids: evolutionary implications for human brain development. World Rev Nutr Diet 2001, 90:144-161.
17. Bravata DM, Sanders L, Huang J, Krumholz HM, Olkin I, Gardner CD, Bravata DM. Efficacy and safety of low-carbohydrate diets: a systematic review.
JAMA. 2003 Apr 9;289(14):1837-50.
18. St Jeor ST, Howard BV, Prewitt TE, Bovee V, Bazzarre T, Eckel RH et al.. Dietary protein and weight reduction: a statement for healthcare professionals from the Nutrition Committee of the Council on Nutrition, Physical Activity, and Metabolism of the American Heart Association. Circulation. 2001 Oct 9;104(15):1869-74
19. Wright JD, J Kennedy-Stephenson J, Wang CY, McDowell MA, Johnson CL, National Center for Health Statistics, CDC. Trends in intake of energy and macronutrients—United States, 1971-2000. JAMA. 2004;291:1193-1194.
20. McDowell M, Briefel R, Alaimo K, et al. Energy and macronutrient intakes of persons ages 2 months and over in the United States: Third National Health and Nutrition Examination Survey, Phase 1, 1988–91.Washington, DC: US Government Printing Office, Vital and Health Statistics; 1994. CDC publication No. 255.
21. Rudman D, DiFulco TJ, Galambos JT, Smith RB 3rd, Salam AA, Warren WD. Maximal rates of excretion and synthesis of urea in normal and cirrhotic subjects.
J Clin Invest. 1973 Sep;52(9):2241-9.
22. Speth JD, Spielmann KA. Energy source, protein metabolism, and hunter-gatherer subsistence strategies. J Anthropological Archaeology 1983;2:1-31.
23. Speth JD. Early hominid hunting and scavenging: the role of meat as an energy source. J Hum Evol 1989;18:329-43.
24. Noli D, Avery G. Protein poisoning and coastal subsistence. J Archaeological Sci 1988;15:395-401.
25. Lieb CW. The effects on human beings of a twelve months’ exclusive meat diet. JAMA 1929;93:20-22.
26. Ogden CL, Fryar CD, Carroll MD, Flegal KM. Mean body weight, height and body mass index, United States 1960—2002 . Center for Disease Control. Advance Data from Vital and Health Statistics, No. 347, October 27, 2004.
27. Oliver WJ, Cohen EL, Neel JV. Blood pressure, sodium intake, and sodium related hormones in the Yanomamo Indians, a “no-salt” culture. Circulation. 1975 Jul;52(1):146-51.
28. Semaw S, Rogers MJ, Quade J, Renne PR, Butler RF, Dominguez-Rodrigo M, Stout D, Hart WS, Pickering T, Simpson SW. 2.6-Million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar, Ethiopia. J Hum Evol. 2003 Aug;45(2):169-77.
29. Bunn, HT, Kroll EM. Systematic butchery by Plio-Pleistocene hominids at Olduvai Gorge, Tanzania. Curr Anthropol 1986;20:365–398.
30. de Heinzelin J, Clark JD, White T, Hart W, Renne P, WoldeGabriel G, Beyene Y, Vrba E. Environment and behavior of 2.5-million-year-old Bouri hominids.
Science. 1999 Apr 23;284(5414):625-9
31. Asfaw B. White T, Lovejoy O, Latimer B, Simpson S, Suwa, G. Australopithecus garhi: A new species of early hominid from Ethiopia. Science 1999; 284, 629–635.
32. Vekua A, Lordkipanidze D, Rightmire GP, Agusti J, Ferring R, Maisuradze G, Mouskhelishvili A, Nioradze M, De Leon MP, Tappen M, Tvalchrelidze M, Zollikofer C. A new skull of early Homo from Dmanisi, Georgia. Science. 2002 Jul 5;297(5578):85-9.
33. Zhu RX, Potts R, Xie F. Hoffman KA, Deng CL, Shi CD, Pan YX, Wang HQ, Shi, RP, Wang YC, Shi GH, Wu NQ. New evidence on the earliest human presence at high northern latitudes in northeast Asia. Nature 2004; 431: 559–562.
34. Aiello LC, Wheeler P. The expensive tissue hypothesis. Curr Anthropol 1995; 36:199–222.
35. Leonard W.R, Robertson ML. Evolutionary perspectives on human nutrition: The influence of brain and body size on diet and metabolism. Am J Hum Biol 1994; 6: 77–88.
36. Pawlosky R., Barnes A., Salem, N. Essential fatty acid metabolism in the feline: Relationship between liver and brain production of long-chain polyunsaturated fatty acids. J Lipid Res 1994;35: 2032–2040.
37. Hussein N, Ah-Sing E, Wilkinson P, Leach C, Griffin BA, Millward DJ. Long-chain conversion of [13C] linoleic acid and alpha-linolenic acid in response to marked changes in their dietary intake in men. J Lipid Res. 2005 Feb;46(2):269-80.
38. Sturman JA, Hepner GW, Hofmann AF, Thomas PJ. Metabolism of [35S] taurine in man. J Nutr. 1975 Sep;105(9):1206-14.
39. Chesney RW, Helms RA, Christensen M, Budreau AM, Han X, Sturman JA. The role of taurine in infant nutrition. Adv Exp Med Biol. 1998;442:463-76.
40. Laidlaw SA, Shultz TD, Cecchino JT, Kopple JD. Plasma and urine taurine levels in vegans. Am J Clin Nutr. 1988 Apr;47(4):660-3.
41. Knopf K, Sturman JA, Armstrong M, Hayes KC. 1978. Taurine: An essential nutrient for the cat. J Nutr 1978;108: 773–778.
42. MacDonald ML, Rogers QR, Morris JG. Nutrition of the domestic cat, a mammalian carnivore. Annu Rev Nutr 1984; 4: 521–562.
43. Fam AG. Gout: excess calories, purines, and alcohol intake and beyond. Response to a urate-lowering diet. J Rheumatol. 2005 May;32(5):773-7.
44. Matzkies F, Berg G, Madl H. The uricosuric action of protein in man. Adv Exp Med Biol 1980;122A:227-31.
45. Loffler W. Grobner W, Medina R, Zollner N. Influence of dietary purines on pool size, turnover, and excretion of uric acid during balance conditions. Isotope studies using 15N-uric acid. Res Exp Med (Berl). 1982(2):113-123.
46. Oda M, Satta Y, Takenaka O, Takahata N. Loss of urate oxidase activity in hominoids and its evolutionary implications. Mol Biol Evol. 2002 May; 19(5): 640-53.
47. Abadeh S, Killacky J, Benboubetra M, Harrison R. Purification and partial characterization of xanthine oxidase from human milk. Biochim Biophys Acta. 1992 Jul 21;1117(1):25-32
48. Xu P, LaVallee P, Hoidal JR. Repressed expression of the human xanthine oxidoreductase gene. E-box and TATA-like elements restrict ground state transcriptional activity. J Biol Chem. 2000 Feb 25;275(8):5918-26.
49. Dessein PH, Shipton EA, Stanwix AE, Joffe BI, Ramokgadi J. Beneficial effects of weight loss associated with moderate calorie/carbohydrate restriction, and increased proportional intake of protein and unsaturated fat on serum urate and lipoprotein levels in gout: a pilot study. Ann Rheum Dis. 2000 Jul;59(7):539-43.
50. Teleki G. The omnivorous chimpanzee. Sci Am 1973; 228: 33–42.
51. Stanford CB. The hunting ecology of wild chimpanzees: Implications for the evolutionary ecology of Pliocene hominids. Am Anthropol 1996; 98: 96–113.
52. Schoeninger MJ, Moore J, Sept JM. Subsistence strategies of two “savanna” chimpanzee populations: The stable isotope evidence. Am J Primatol 1999: 49: 297–314.
53. van der Merwe NJ, Thackeray JF, Lee-Thorp JA, Luyt J. The carbon isotope ecology and diet of Australopithecus africanus at Sterkfontein, South Africa J Hum Evol 2003;44: 581–597.
54. Lee-Thorp J, Thackeray JF, van der Merwe N. The hunters and the hunted revisited. J Hum Evol 2000; 39: 565–576.
55. Sponheimer M, Lee-Thorp JA.. Differential resource utilization by extant great apes and australopithecines: Towards solving the C4 conundrum. Comp Biochem Physiol A 2003;136: 27–34.
56. Sussman RW. Foraging patterns of nonhuman primates and the nature of food preferences in man. Fed Proc 1978;37: 55–60.
57. Richards MP, Pettitt PB, Trinkaus E, Smith FH, Paunovic M, Karavanic, I. Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes. Proc Natl Acad Sci 2000;97: 7663–7666.
58. Bocherens H, Drucker DG, Billiou D, Patou-Mathis M, Vandermeersch B. Isotopic evidence for diet and subsistence pattern of the Saint-Cesaire I Neanderthal: review and use of a multi-source mixing model. J Hum Evol. 2005 Jul;49(1):71-87
59. Balter V, Simon L. Diet and behavior of the Saint-Cesaire Neanderthal inferred from biogeochemical data inversion. J Hum Evol. 2006 Oct;51(4):329-38.
60. Currat M, Excoffier L. Modern humans did not admix with Neanderthals during their range expansion into Europe. PLoS Biol. 2004 Dec;2(12):e421. Epub 2004 Nov 30
61. Richards MP, Hedges REM, Jacobi R, Current, A, Stringer C. Focus: Gough’s Cave and Sun Hole Cave human stable isotope values indicate a high animal protein diet in the British Upper Palaeolithic. J Archaeol Sci 2000;27: 1–3.
62. Gray JP. A corrected ethnographic atlas. World Cult J 1999;10: 24–85.
63. Hayden B. Subsistence and ecological adaptations of modern hunter/gatherers. In: RSO Harding, RSO, Teleki G. (Eds.), Omnivorous Primates. Columbia University Press, New York, 1981, pp. 344–421.
64. Kaplan H, Hill K, Lancaster J, Hurtado AM. A theory of human life history evolution: diet, intelligence, and longevity. Evol. Anthropol 2000;9:156–185.
65. Sempos CT, Liu K, Ernst ND. Food and nutrient exposures: what to consider when evaluating epidemiologic evidence. Am J Clin Nutr. 1999 Jun;69(6):1330S-1338S.
66. Potischman N, Weed DL. Causal criteria in nutritional epidemiology.
Am J Clin Nutr. 1999 Jun;69(6):1309S-1314S.
67. Freudenheim JL. Study design and hypothesis testing: issues in the evaluation of evidence from research in nutritional epidemiology. Am J Clin Nutr. 1999 Jun; 69(6): 1315S-1321S.
68. Fraser GE. A search for truth in dietary epidemiology. Am J Clin Nutr. 2003 Sep;78(3 Suppl):521S-525S.
69. Flegal KM. Evaluating epidemiologic evidence of the effects of food and nutrient exposures. Am J Clin Nutr. 1999 Jun;69(6):1339S-1344S.
70. Hu FB, Stampfer MJ, Manson JE, Rimm E, Colditz GA, Speizer FE, Hennekens CH, Willett WC. Dietary protein and risk of ischemic heart disease in women.
Am J Clin Nutr. 1999 Aug;70(2):221-7.
71. Campbell TC, Junshi C. Diet and chronic degenerative diseases: perspectives from China. Am J Clin Nutr. 1994 May;59(5 Suppl):1153S-1161S.
72. Campbell TC, Parpia B, Chen J. Diet, lifestyle, and the etiology of coronary artery disease: the Cornell China study. Am J Cardiol. 1998 Nov 26;82(10B):18T-21T.
73. Popkin BM. Where’s the fat? Trends in U.S. Diets 1965-1996. Prev Med 2001;32:245-54.
74. O’Dea K, Traianedes K, Chisholm K, Leyden H, Sinclair AJ. Cholesterol-lowering effect of a low-fat diet containing lean beef is reversed by the addition of beef fat. Am J Clin Nutr. 1990 Sep;52(3):491-4.
75. Wolfe BM & Giovannetti PM (1991): Short term effects of substituting protein for carbohydrate in the diets of moderately hypercholesterolemic human subjects. Metabolism 40, 338-343.
76. Wolfe BM & Piche LA (1999): Replacement of carbohydrate by protein in a conventional-fat diet reduces cholesterol and triglyceride concentrations in healthy normolipidemic subjects. Clin. Invest. Med. 22, 140-148.
77. O’Dea K (1984): Marked improvement in carbohydrate and lipid metabolism in diabetic Australian Aborigines after temporary reversion to traditional lifestyle. Diabetes 33, 596-603.
78. O’Dea K, Traianedes K, Ireland P, Niall M, Sadler J, Hopper J & DeLuise M (1989): The effects of diet differing in fat, carbohydrate, and fiber on carbohydrate and lipid metabolism in type II diabetes. J. Am. Diet. Assoc. 89, 1076-1086.
79. Layman DK, Boileau RA, Erickson DJ, Painter JE, Shiue H, Sather C, Christou DD. A reduced ratio of dietary carbohydrate to protein improves body composition and blood lipid profiles during weight loss in adult women. J Nutr. 2003 Feb;133(2):411-7.
80. Noakes M, Keogh JB, Foster PR, Clifton PM. Effect of an energy-restricted, high-protein, low-fat diet relative to a conventional high-carbohydrate, low-fat diet on weight loss, body composition, nutritional status, and markers of cardiovascular health in obese women. Am J Clin Nutr. 2005 Jun;81(6):1298-306.
81. Farnsworth E, Luscombe ND, Noakes M, Wittert G, Argyiou E, Clifton PM. Effect of a high-protein, energy-restricted diet on body composition, glycemic control, and lipid concentrations in overweight and obese hyperinsulinemic men and women.
Am J Clin Nutr. 2003 Jul;78(1):31-9.
82. Luscombe-Marsh ND, Noakes M, Wittert GA, Keogh JB, Foster P, Clifton PM.
Carbohydrate-restricted diets high in either monounsaturated fat or protein are equally effective at promoting fat loss and improving blood lipids. Am J Clin Nutr. 2005 Apr;81(4):762-72
83. Aude YW, Agatston AS, Lopez-Jimenez F, Lieberman EH, Marie Almon, Hansen M, Rojas G, Lamas GA, Hennekens CH. The national cholesterol education program diet vs a diet lower in carbohydrates and higher in protein and monounsaturated fat: a randomized trial. Arch Intern Med. 2004 Oct 25;164(19):2141-6.
84. McAuley KA, Hopkins CM, Smith KJ, McLay RT, Williams SM, Taylor RW, Mann JI. Comparison of high-fat and high-protein diets with a high-carbohydrate diet in insulin-resistant obese women. Diabetologia. 2005 Jan;48(1):8-16.
85. Appel LJ, Sacks FM, Carey VJ, Obarzanek E, Swain JF, Miller ER 3rd, Conlin PR, Erlinger TP, Rosner BA, Laranjo NM, Charleston J, McCarron P, Bishop LM; OmniHeart Collaborative Research Group. Effects of protein, monounsaturated fat, and carbohydrate intake on blood pressure and serum lipids: results of the OmniHeart randomized trial. JAMA. 2005 Nov 16;294(19):2455-64.
86. Kalopissis, AD Griffaton G, Fau D. Inhibition of hepatic very low density lipoprotein secretion on obese Zucker rats adapted to a high protein diet. Metabolism 1995;44:19-29.
87. Appel LJ. The effects of protein intake on blood pressure and cardiovascular disease. Curr Opin Lipidol. 2003 Feb;14(1):55-9. Review.
88. Elliott P. Protein intake and blood pressure in cardiovascular disease.
Proc Nutr Soc. 2003 May;62(2):495-504.
89. He J, Whelton PK. Elevated systolic blood pressure as a risk factor for cardiovascular and renal disease. J Hypertens Suppl. 1999 Jun;17(2):S7-13.
90. Burke V, Hodgson JM, Beilin LJ, Giangiulioi N, Rogers P, Puddey IB.
Dietary protein and soluble fiber reduce ambulatory blood pressure in treated hypertensives. Hypertension. 2001 Oct;38(4):821-6.
91. Washburn S, Burke GL, Morgan T, Anthony M. Effect of soy protein supplementation on serum lipoproteins, blood pressure, and menopausal symptoms in perimenopausal women. Menopause. 1999 Spring;6(1):7-13.
92. He J, Gu D, Wu X, Chen J, Duan X, Chen J, Whelton PK. Effect of soybean protein on blood pressure: a randomized, controlled trial.Ann Intern Med. 2005 Jul 5;143(1):1-9.
93. Hodgson JM, Burke V, Beilin LJ, Puddey IB. Partial substitution of carbohydrate intake with protein intake from lean red meat lowers blood pressure in hypertensive persons. Am J Clin Nutr. 2006 Apr;83(4):780-7
94. Nuttall FQ, Gannon MC. The metabolic response to a high-protein, low-carbohydrate diet in men with type 2 diabetes mellitus. Metabolism. 2006 Feb;55(2):243-51.
95. Nuttall FQ, Gannon MC. Metabolic response of people with type 2 diabetes to a high protein diet. Nutr Metab (Lond). 2004 Sep 13;1(1):6
52.
96. McAuley KA, Smith KJ, Taylor RW, McLay RT, Williams SM, Mann JI. Long-term effects of popular dietary approaches on weight loss and features of insulin resistance. Int J Obes (Lond). 2006 Feb;30(2):342-9.
97. Weigle DS, Breen PA, Matthys CC, Callahan HS, Meeuws KE, Burden VR, Purnell JQ. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am J Clin Nutr. 2005 Jul;82(1):41-8
98. Due A, Toubro S, Skov AR, Astrup A. Effect of normal-fat diets, either medium or high in protein, on body weight in overweight subjects: a randomised 1-year trial.
Int J Obes Relat Metab Disord. 2004 Oct;28(10):1283-90
99. Westerterp-Plantenga MS, Lejeune MP, Nijs I, van Ooijen M, Kovacs EM. High protein intake sustains weight maintenance after body weight loss in humans. Int J Obes Relat Metab Disord. 2004 Jan;28(1):57-64.
100. Lejeune MP, Kovacs EM, Westerterp-Plantenga MS. Additional protein intake limits weight regain after weight loss in humans. Br J Nutr. 2005 Feb;93(2):281-9.
101. Porrini M, Santangelo A, Crovetti R, Riso P, Testolin G, Blundell JE.
Reid M, Hetherington M. Relative effects of carbohydrates and protein on satiety -- a review of methodology. Neurosci Biobehav Rev 1997 May;21(3):295-308
102. Poppitt SD, McCormack D, Buffenstein R. Short-term effects of macronutrient preloads on appetite and energy intake in lean women. Physiol Behav 1998 Jun 1;64(3):279-85
103. Batterham RL, Heffron H, Kapoor S, Chivers JE, Chandarana K, Herzog H, Le Roux CW, Thomas EL, Bell JD, Withers DJ. Critical role for peptide YY in protein-mediated satiation and body-weight regulation. Cell Metab. 2006 Sep;4(3):223-33.
104. O’Brien KO, Abrams SA, Liang LK, Ellis KJ, Gagel RF. Increased efficiency of calcium absorption during short periods of inadequate calcium intake in girls. Am J Clin Nutr. 1996 Apr;63(4):579-83
105. Dawson-Hughes B, Harris SS, Rasmussen H, Song L, Dallal GE. Effect of dietary protein supplements on calcium excretion in healthy older men and women. J Clin Endocrinol Metab. 2004 Mar;89(3):1169-73
106. Spencer H, Kramer L, Osis D, Norris C. Effect of a high protein (meat) intake on calcium metabolism in man. Am J Clin Nutr. 1978 Dec;31(12):2167-80
107. Roughead ZK, Johnson LK, Lykken GI, Hunt JR. Controlled high meat diets do not affect calcium retention or indices of bone status in healthy postmenopausal women.
J Nutr. 2003 Apr;133(4):1020-6
108. Arjmandi BH, Khalil DA, Smith BJ, Lucas EA, Juma S, Payton ME, Wild RA.
Soy protein has a greater effect on bone in postmenopausal women not on hormone replacement therapy, as evidenced by reducing bone resorption and urinary calcium excretion. J Clin Endocrinol Metab. 2003 Mar;88(3):1048-54
109. Pannemans DL, Schaafsma G, Westerterp KR. Calcium excretion, apparent calcium absorption and calcium balance in young and elderly subjects: influence of protein intake. Br J Nutr. 1997 May;77(5):721-9.
110. Kerstetter JE, O’Brien KO, Insogna KL. Dietary protein affects intestinal calcium absorption. Am J Clin Nutr. 1998 Oct;68(4):859-65.
111. Kerstetter JE, O’Brien KO, Caseria DM, Wall DE, Insogna KL. The impact of dietary protein on calcium absorption and kinetic measures of bone turnover in women.
J Clin Endocrinol Metab. 2005 Jan;90(1):26-31.
112. Kerstetter JE, Wall DE, O’Brien KO, Caseria DM, Insogna KL. Meat and soy protein affect calcium homeostasis in healthy women. J Nutr. 2006 Jul;136(7):1890-5
113. Brenner BM, Meyer TW, Hostetter TH. Dietary protein intake and the progressive nature of kidney disease: the role of hemodynamically mediated glomerular injury in the pathogenesis of progressive glomerular sclerosis in aging, renal ablation, and intrinsic renal disease.N Engl J Med. 1982 Sep 9;307(11):652-9.
114. Lippert J, Ritz E, Schwarzbeck A, Schneider P. The rising tide of endstage renal failure from diabetic nephropathy type II--an epidemiological analysis.
Nephrol Dial Transplant. 1995;10(4):462-7.
115. Ritz E, Rychlik I, Locatelli F, Halimi S. End-stage renal failure in type 2 diabetes: A medical catastrophe of worldwide dimensions. Am J Kidney Dis. 1999 Nov;34(5):795-808.
116. Wrone EM, Carnethon MR, Palaniappan L, Fortmann SP; Third National Health and Nutrition Examination Survey. Association of dietary protein intake and microalbuminuria in healthy adults: Third National Health and Nutrition Examination Survey. Am J Kidney Dis. 2003 Mar;41(3):580-7.
117. Johnson DW. Dietary protein restriction as a treatment for slowing chronic kidney disease progression: the case against. Nephrology. 2006 Feb;11(1):58-62.
118. Skov AR, Toubro S, Bulow J, Krabbe K, Parving HH, Astrup A. Changes in renal function during weight loss induced by high vs low-protein low-fat diets in overweight subjects. Int J Obes Relat Metab Disord. 1999 Nov;23(11):1170-7.
119. Bingham SA. High-meat diets and cancer risk. Proc Nutr Soc. 1999 May;58(2):243-8.
120. Truswell AS. Meat consumption and cancer of the large bowel. Eur J Clin Nutr. 2002 Mar;56 Suppl 1:S19-24.
121. Larsson SC, Wolk A. Meat consumption and risk of colorectal cancer: A meta-analysis of prospective studies.Int J Cancer. 2006 Sep 21; [Epub ahead of print]
122. Bingham SA, Hughes R, Cross AJ. Effect of white versus red meat on endogenous N-nitrosation in the human colon and further evidence of a dose response. J Nutr. 2002 Nov;132(11 Suppl):3522S-3525S
123. Lunn JC, Kuhnle G, Mai V, Frankenfeld C, Shuker DE, Glen RC, Goodman JM, Pollock JR, Bingham SA. The effect of haem in red and processed meat on the endogenous formation of N-nitroso compounds in the upper gastrointestinal tract. Carcinogenesis. 2006 Oct 19; [Epub ahead of print].
124. Haorah J, Zhou L, Wang X, Xu G, Mirvish SS. Determination of total N-nitroso compounds and their precursors in frankfurters, fresh meat, dried salted fish, sauces, tobacco, and tobacco smoke particulates. J Agric Food Chem. 2001 Dec;49(12):6068-78
125. Gerrior S, Bente I. 2002. Nutrient Content of the U.S. Food Supply, 1909-99: A Summary Report.
U.S.D.A, Center for Nutrition Policy and Promotion. Home Economics Research Report No. 55.
126. Hughes R, Pollock JR, Bingham S. Effect of vegetables, tea, and soy on endogenous N-nitrosation, fecal ammonia, and fecal water genotoxicity during a high red meat diet in humans. Nutr Cancer. 2002;42(1):70-7.
127. Stefansson V. Cancer: Disease of Civilization. Hill and Wang, New York, 1960.
128. Patnaik AK, Liu SK, Johnson GF. Feline intestinal adenocarcinoma. A clinicopathologic study of 22 cases. Vet Pathol. 1976;13(1):1-10
129. Anthony JC, Lang CH, Crozier SJ, Anthony TG, MacLean DA, Kimball SR, Jefferson LS. Contribution of insulin to the translational control of protein synthesis in skeletal muscle by leucine. Am J Physiol Endocrinol Metab. 2002;282:E1092-101.
130. Layman DK. Role of leucine in protein metabolism during exercise and recovery. Can J Appl Physiol 2002;27:646-62.
131. Rasmussen BB, Tipton KD, Miller SL, Wolf SE, Wolfe RR. An oral essential amino acid-carbohydrate supplement enhances muscle protein anabolism after resistance exercise.J Appl Physiol. 2000 Feb;88(2):386-92.
132. Tipton KD, Rasmussen BB, Miller SL, Wolf SE, Owens-Stovall SK, Petrini BE, Wolfe RR. Timing of amino acid-carbohydrate ingestion alters anabolic response of muscle to resistance exercise. Am J Physiol Endocrinol Metab. 2001 Aug;281(2):E197-206.
133. Levenhagen DK, Gresham JD, Carlson MG, Maron DJ, Borel MJ, Flakoll PJ. Postexercise nutrient intake timing in humans is critical to recovery of leg glucose and protein homeostasis. Am J Physiol Endocrinol Metab. 2001;280:E982-93.
134. Blomstrand E. Amino acids and central fatigue. Amino Acids 2001;20:25-34.
135. Kuhn T. The Structure of Scientific Revolutions. University of Chicago Press, Chicago, 1970.
136. Nishikimi M, Yagi K. Molecular basis for the deficiency in humans of gulonolactone oxidase, a key enzyme for ascorbic acid biosynthesis. Am J Clin Nutr. 1991 Dec;54(6 Suppl):1203S-1208S.
137. Rand WM, Pellett PL, Young VR. Meta-analysis of nitrogen balance studies for estimating protein requirements in healthy adults. Am J Clin Nutr. 2003 Jan;77(1):109-27.
138. Vieth R. Why the optimal requirement for Vitamin D3 is probably much higher than what is officially recommended for adults. J Steroid Biochem Mol Biol. 2004 May;89-90(1-5):575-9.
139. Dawson-Hughes B, Heaney RP, Holick MF, Lips P, Meunier PJ, Vieth R. Estimates of optimal vitamin D status. Osteoporos Int. 2005 Jul;16(7):713-6.
140. Miller M. Niacin as a component of combination therapy for dyslipidemia.
Mayo Clin Proc. 2003 Jun;78(6):735-42.
141. Chittenden RH. Physiologic economy in nutrition. New York: Heinemann, 1905.
142. Remer T, Manz F. Potential renal acid load of foods and its influence on urine pH. J Am Diet Assoc. 1995 Jul;95(7):791-7.
143. Cordain L Cereal grains: humanity’s double-edged sword. World Rev Nutr Diet. 1999;84:19-73.
144. Berlyne GM, Ben Ari J, Nord E, Shainkin R: Bedouin osteomalacia due to calcium deprivation caused by high phytic acid content of unleavened bread. Am J Clin Nutr 1973;26:910-11.
145. Ford JA, Colhoun EM, McIntosh WB, Dunnigan MG: Biochemical response of late rickets and osteomalacia to a chupatty-free diet. Brit Med J 1972;2:446-47.
146. Robertson I, Ford JA, McIntosh WB, Dunnigan MG: The role of cereals in the aetiology of nutritional rickets: the lesson of the Irish national nutritional survey 1943-8. Brit J Nutr 1981;45:17-22.
147. Stephens WP, Berry JL, Klimiuk PS, Mawer EB: Annual high dose vitamin D prophylaxis in asian immigrants. Lancet 1981;2:1199-201.
148. Ewer TK: Rachitogenicity of green oats. Nature 1950;166:732-33.
149. MacAuliffe T, Pietraszek A, McGinnis J: Variable rachitogenic effects of grain and alleviation by extraction or supplementation with vitamin D, fat and antibiotics. Poultry Sci 1976;55:2142-47.
150. Hidiroglou M, Ivan M, Proulx JG, Lessard JR: Effect of a single intramuscular dose of vitamin D on concentrations of liposoluble vitamins in the plasma of heifers winter-fed oat silage, grass silage or hay. Can J Anim Sci 1980;60:311-18.
151. Sly MR, van der Walt WH, Du Bruyn DB, Pettifor JM, Marie PJ: Exacerbation of rickets and osteomalacia by maize: a study of bone histomorphometry and composition in young baboons. Calcif Tissue Int 1984;36:370-79.
152. Batchelor AJ, Compston JE: Reduced plasma half-life of radio-labelled 25-hydroxyvitamin D3 in subjects receiving a high fiber diet. Brit J Nutr 1983;49:213-16.
153. Clements MR, Johnson L, Fraser DR: A new mechanism for induced vitamin D deficiency in calcium deprivation. Nature 1987;325:62-65.
154. Navarro A, Diaz MP, Munoz SE, Lantieri MJ, Eynard AR. Characterization of meat consumption and risk of colorectal cancer in Cordoba, Argentina.Nutrition. 2003 Jan;19(1):7-10.
155. Muscat JE, Wynder EL. The consumption of well-done red meat and the risk of colorectal cancer. Am J Public Health. 1994 May;84(5):856-8.
156. Sinha R, Chow WH, Kulldorff M, Denobile J, Butler J, Garcia-Closas M, Weil R, Hoover RN, Rothman N. Well-done, grilled red meat increases the risk of colorectal adenomas. Cancer Res. 1999 Sep 1;59(17):4320-4
157. Williams JH, Phillips TD, Jolly PE, Stiles JK, Jolly CM, Aggarwal D Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. Am J Clin Nutr. 2004 Nov;80(5):1106-22.
158. Dunaif GE, Campbell TC. Dietary protein level and aflatoxin B1-induced preneoplastic hepatic lesions in the rat. J Nutr. 1987 Jul;117(7):1298-302.
159. Dunaif GE, Campbell TC. Relative contribution of dietary protein level and aflatoxin B1 dose in generation of presumptive preneoplastic foci in rat liver.
J Natl Cancer Inst. 1987 Feb;78(2):365-9.
160. Horio F, Youngman LD, Bell RC, Campbell TC. Thermogenesis, low-protein diets, and decreased development of AFB1-induced preneoplastic foci in rat liver.
Nutr Cancer. 1991;16(1):31-41.
161. Youngman LD, Campbell TC. The sustained development of preneoplastic lesions depends on high protein intake. Nutr Cancer. 1992;18(2):131-42.
162. Fasano A, Berti I, Gerarduzzi T, Not T, Colletti RB, Drago S, Elitsur Y, Green PH, Guandalini S, Hill ID, Pietzak M, Ventura A, Thorpe M, Kryszak D, Fornaroli F, Wasserman SS, Murray JA, Horvath K. Prevalence of celiac disease in at-risk and not-at-risk groups in the United States: a large multicenter study.Arch Intern Med. 2003 Feb 10;163(3):286-92.
163. Cordain L, Toohey L, Smith MJ, Hickey MS. Modulation of immune function by dietary lectins in rheumatoid arthritis. Br J Nutr. 2000 Mar;83(3):207-17.
164. Key TJ, Fraser GE, Thorogood M, Appleby PN, Beral V, Reeves G, Burr ML, Chang-Claude J, Frentzel-Beyme R, Kuzma JW, Mann J, McPherson K. Mortality in vegetarians and nonvegetarians: detailed findings from a collaborative analysis of 5 prospective studies. Am J Clin Nutr. 1999 Sep;70(3 Suppl):516S-524S


